Лекция1.Введение.Предметизадачикурса.Зоологическаяклассифика-ция.ЦарствоПротисты.ТипРизоподы.Общаяхарактеристика.КлассЛобозеи.
реклама

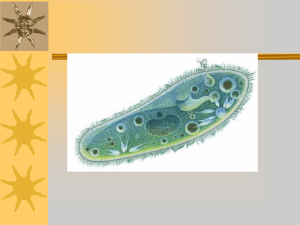
1 Лекция 1. Тема лекции: Введение в зоологию беспозвоночных План занятий: Предмет и задачи курса. Зоологическая классификация. Царство Протисты. Тип Ризоподы. Общая характеристика. Класс Лобозеи. Начало накопления человеком сведений о животном мире относится к каменному веку (палеолиту). Животные были объектом охоты, рыболовства и т. п. Сочинения о животных известны уже в Древнем Китае, Индии. Широко распространены изображения животных, в том числе и беспозвоночных, в которых наивная фантазия нередко сочетается с точными и живыми деталями. Однако научная зоология берет начало от великого ученого и мыслителя Древней Греции Аристотеля (IV в. до н. э.). Он разделил всех известных ему животных (их было около 500) на две группы: 1) животные, имеющие кровь, и 2) животные без крови. К первой группе он отнес всех высших животных (зверей, птиц, гадов и рыб), ко второй — насекомых, раков, моллюсков и других низших животных. Эта первая зоологическая система просуществовала очень долго. Кроме того, в работах Аристотеля высказывается ряд важных идей и обобщений, в том числе и учение о корреляциях частей организма. Средневековый феодализм с характерным для него почти безграничным господством церкви подавлял движение научной мысли. Лишь в XIV в., в эпоху Возрождения, начинается развитие естествознания вообще и зоологии в частности. В течение XVI — XVII вв. происходит первоначальное накопление сведений о многообразии животных, их строении, образе жизни (сочинения К. Геснера в Швейцарии, Г. Ронделе и П. Белона во Франции и др.). Большое значение для развития зоологии на рубеже XVI - XVII вв. имело изобретение микроскопа, положившее начало познанию нового мира самых мелких живых существ, исследованию тонкого строения организмов и их эмбрионального развития (А. Левенгук в Голландии, Мальпиги в Италии, У. Гарвей в Англии и др.). В конце XVII и в первой половине XVIII в. закладываются основы системы животного мира. Большое значение в этом плане имели работы Дж. Рея (Англия) и в особенности выдающегося шведского естествоиспытателя К. Линнея, который ввел рациональную номенклатуру, сыгравшую важную роль в развитии систематической зоологии и ботаники. Его классический труд «Systema naturae» («Система природы») впервые вышел в 1735 г., а в 10-м издании его (1758) уже последовательно были разработаны принципы 2 бинарной номенклатуры. В системе Линнея различались 4 взаимно подчиненные систематические категории-таксоны: вид, род, порядок, класс. Он установил и назвал более 300 родов животных, которые по степени сходства сгруппировал в порядки. Сходные порядки были объединены в классы, которые рассматривались как высшие систематические категории. Линней установил 6 классов: 1. Mammalia (млекопитающие); 2. Aves (птицы); 3. Amphibia (гады); 4. Pisces (рыбы); 5. Insecta (насекомые) и 6. Vermes (черви, моллюски и все прочие низшие животные). К. Линней стоял на позициях учения о неизменяемости видов. В конце XVIII и начале XIX в. французский зоолог Ж. Кювье разработал основы сравнительной анатомии животных и, в частности, учение о корреляциях. На основе этих работ его ученик Бленвиль в 1825 г. вводит в систему понятие тип как высшую таксономическую единицу. Сначала таких типов было установлено немного. Потом с углублением знаний о строении и развитии животных количество их увеличилось. Ж. Кювье, как и К. Линней, считал виды неизменяющимися. В первой половине XIX в. в зоологии появляется идея исторического развития животного мира. Современник и соотечественник Ж. Кювье, Э. Жоффруа Сент Илер развивал идею изменяемости видов под прямым воздействием факторов среды. В этот же период Ж. Б. Ламарк опубликовал книгу «Философия зоологии» (1809), в которой излагалась первая научная теория эволюции органического мира. Ламарк много сделал также и для разработки системы беспозвоночных животных. Ему принадлежит термин «беспозвоночные», среди которых он различал 10 классов (у Линнея было лишь 2 класса). Против идеи неизменяемости видов в этот же период в России выступил профессор Московского университета К. Ф. Рулье. Большую роль в развитии зоологии в середине XIX в. сыграл академик Российской Академии Наук К. М. Бэр, автор выдающихся исследований в области эмбриологии животных, создатель учения о зародышевых листках. Большое влияние на развитие зоологии оказала сформулированная в конце 30-х годов XIX в. клеточная теория, созданная трудами М. Шлейдена и Т. Шванна. Эти работы убедительно показали единство микроскопической структуры животных и растений. Новый период в развитии зоологии, как и всех биологических наук, начинается во второй половине XIX в. после работ Ч. Дарвина, утвердившего в своем знаменитом труде «Происхождение видов путем естественного отбора, или сохранение благоприятствуемых пород в борьбе за жизнь» (1859) эволюционное учение и открывшего основной фактор органической эволюции — естественный отбор. На основе эволюционного учения зоология стала быстро развиваться, и возникли новые, ранее не существовавшие зоологические дисциплины. В Германии Э. Геккель использует идеи Ч. Дарвина для разработки филогении животного мира. Ему принадлежит и классическая формулировка «основного биогенетического закона», устанавливающего определенное соотношение между онтогенезом (индивидуальным развитием) и филогенезом 3 (историческим развитием). Честь открытия этих закономерностей принадлежит крупному зоологу и эмбриологу Ф. Мюллеру. Возникают эволюционная сравнительная анатомия (Р. Видерсгейм, К. Гегенбаур, Э. Рей Ланкастер и др.) и эволюционная сравнительная эмбриология. В создании последней ведущая роль принадлежит русским ученым, в первую очередь И. И. Мечникову и А. О. Ковалевскому. В этот же период В. О. Ковалевский исследованиями по ископаемым копытным закладывает основы эволюционной палеозоологии (наука, изучающая ископаемых животных). Быстрыми темпами происходит развитие систематики и зоогеографии животных. Об этом свидетельствует число описанных видов. К. Линиею было известно 4208 видов, в первой половине XIX в. это число возросло до 48000, а в конце века зоологи насчитывали около 400 тыс. видов. В ХХ в. (и до наших дней) продолжается работа зоологов по развитию и усовершенствованию системы животного мира. На основе этих исследований значительно возросло количество высших систематических категорий — типов и классов. Во времена Кювье различали 4 типа, в современных же системах их насчитывается много больше. Результаты зоологических исследований в ХХ в. характеризуются значительными достижениями в области филогенетики — познании конкретных путей эволюции животного мира. В разработке этих проблем важное значение имеют успехи сравнительной анатомии и эмбриологии, а также палеозоологии. Вопросам филогении мы уделим в этой книге существенное внимание. Развитие зоологии в России и Белоруссии неразрывно связано с мировой наукой, но имеет и свои характерные особенности. История развития зоологии тесно связана с историей формирования основных принципов систематики животных. И действительно, разобраться во всем многообразии фауны Земли было бы невозможно, если бы ученые не имели в руках универсального аппарата, позволяющего им точно фиксировать положение изучаемых и описываемых организмов на филогенетическом древе животного царства. Таким аппаратом является современная систематика, возникшая в результате кропотливой работы многих зоологов на протяжении всей истории развития науки. На смену часто весьма формальным построениям прошлого приходит естественная система, в которой ученые стремятся отразить реальные пути эволюции животного мира. В настоящее время царство животных принято делить на серию взаимоподчиненных систематических категорий — таксонов. Основной таксон — «вид». Для обозначения видов используется принцип бинарной номенклатуры, разработанный еще К. Линнеем. Каждому виду присваивается латинское название, состоящее из двух слов. Первое слово — существительное — название рода, в который объединена группа близких видов, второе — обычно прилагательное — название вида. Так, например, научное название белянки капустной — Pieris brassicae, тогда как близкородственные виды, относимые к тому же роду — Pieris, называются: репница — Pieris rapae, брюквенница — Pieris napi и т. д. Двойные названия удобны, так как сразу 4 указывают на родовую принадлежность данного вида. Близкородственные роды объединяются в семейства, семейства - в отряды, отряды - в классы. Высший таксон современной систематики — тип, который объединяет несколько родственных классов. Очень часто зоологами используются «промежуточные» таксоны: подтипы, подклассы, надотряды, подотряды и т. п., объединяющие в пределах данного таксона группы более низкого ранга. Например, классы в пределах типа могут быть сгруппированы в несколько подтипов. Для примера укажем основные систематические категории, к которым причисляется какой-нибудь определенный вид, скажем, белянка капустная. Тип Arthropoda — Членистоногие Подтип Tracheata — Трахейные Надкласс Шестиногие - Hexapoda Класс Насекомые открыточелюстные - Insecta-Ectognatha Отряд Lepidoptera — Чешуекрылые, или Бабочки Семейство Pieridae — Белянки Род Pieris — Белянка Вид Pieris brassicae — Белянка капустная. В современной зоологии, кроме того, явно ощущается необходимость классифицировать и высшие систематические категории — типы. Под одним названием объединяют сходные по существенным признакам родственные типы. Так, всех животных принято делить на царства Одноклеточных (Protista) и Животные (Zoa). Все типы многоклеточных естественно распадаются на три большие группы (надразделы): Фагоцителлообразные (Phagacytellozoa) с одним типом — пластинчатые животные (Placozoa) Низшие многоклеточные (Parazoa) тоже с одним типом — Губки (Spongia) и остальные многоклеточные (Eumetazoa), включающие все оставшиеся типы животных и распадающиеся на Лучистых (Radiata) и Билатеральных многоклеточных (Bilateria). В дальнейшем изложении мы примем следующие подразделения животного царства на группы, стоящие выше типа: царство, подцарство, надраздел, раздел, подраздел. ЦАРСТВО - PROTISTA К одноклеточным (Protista) относятся животные, тело которых морфологически соответствует одной клетке, будучи вместе с тем самостоятельным организмом со всеми присущими организму функциями. Простейшие — это организмы на клеточном уровне организации. В морфологическом отношении тело их равноценно клетке, но в физиологическом представляет целый самостоятельный организм. Подавляющее большинство их микроскопически малых размеров. Общее число известных видов превышает 30000. Строение простейших чрезвычайно разнообразно, но все они обладают чертами, характерными для организации и функции клетки. Два основных 5 компонента тела простейших — цитоплазма и ядро. Цитоплазма ограничена наружной мембраной, которая, как показывает электронный микроскоп, имеет толщину около 7,5 нм и состоит из трех слоев, примерно по 2,5 нм каждый. Эта основная мембрана, состоящая из белков и липоидов и регулирующая поступление веществ в клетку, у многих простейших усложняется дополнительными структурами, увеличивающими толщину и механическую прочность наружного слоя цитоплазмы. Таким образом, возникают образования типа пелликулы и оболочки, которые будут рассмотрены ниже при описании отдельных типов и классов простейших. Цитоплазма простейших обычно распадается на два слоя — наружный, более светлый и плотный, — эктоплазму и внутренний, снабженный многочисленными включениями, — эндоплазму. В цитоплазме локализуются общеклеточные органоиды: митохондрии, эндоплазматическая сеть, рибосомы, элементы аппарата Гольджи. Кроме того, в цитоплазме многих простейших могут присутствовать разнообразные специальные органеллы. Особенно широко распространены различные фибриллярные образования — опорные и сократимые волоконца, сократительные вакуоли, пищеварительные вакуоли и др. Простейшие обладают типичным клеточным ядром, одним или несколькими. Прежние представления о примитивности структуры и формах деления ядра простейших в свете современных данных не соответствуют действительности. Ядро Protozoa обладает типичной двухслойной ядерной оболочкой, пронизанной многочисленными порами. Содержимое ядра: состоит из ядерного сока (кариоплазмы), в котором распределен хроматиновый материал и ядрышки. Жизненный цикл. Клетки, входящие в состав многоклеточного организма (за исключением половых), не имеют жизненного цикла, они дифференцируются, входя в состав той или иной ткани, того или иного органа, и выполняют функции, направленные на сохранение целостности и жизни многоклеточного организма, жизнь многих клеток при этом оказывается кратковременной, они уступают место новым поколениям таких же клеток, как, например, эпителиальные клетки кожи или кишечника. Другие, напротив (нервные клетки), сохраняются в течение всей жизни многоклеточного организма и погибают лишь вместе с ним. В отличие от соматических клеток многоклеточных простейшие характеризуются наличием жизненного цикла. Последний слагается из ряда следующих друг за другом стадий, которые в существовании каждого вида повторяются с определенной закономерностью. Это явление называется цикличностью, а отрезок жизни вида между двумя однозначными стадиями составляет его жизненный цикл. Важная биологическая особенность многих простейших — способность к инцистированию. При этом животные округляются, сбрасывают или втягивают органеллы движения, выделяют на своей поверхности плотную оболочку и переходят от активной жизни в состояние покоя. В инцистированном состоянии простейшие, могут переносить резкие изменения окружающей среды (подсушивание, охлаждение и т. п.), сохраняя жизнеспособность. При возвращении благоприятных для жизни условий цисты раскрываются и 6 простейшие выходят из них в виде активных, подвижных особей. Среды обитания простейших. Простейшие обладают широким всесветным распространением. Множество их живет в море. Некоторые входят в состав бентоса на различных глубинах — от литорали до абиссали (фораминиферы, инфузории, жгутиконосцы). Многочисленные виды радиолярий, жгутиконосцев и инфузорий являются компонентами морского планктона. Многие простейшие (жгутиконосцы, инфузории, корненожки) входят в состав пресноводного бентоса и планктона. Существуют некоторые виды, живущие во влажной почве. Наконец, широкое распространение среди всех классов простейших получил паразитизм. Многие виды паразитических простейших вызывают тяжелые заболевания человека, домашних и промысловых животных. Некоторые виды паразитируют в растениях. Таким образом, простейшие имеют важное практическое значение для медицины, ветеринарии, сельского хозяйства. Классификация Protista. Простейших еще недавно рассматривали как один тип животного мира. В настоящее время, главным образом благодаря исследованиям с электронным микроскопом установлено, что в пределах подцарства Protozoa существуют разные планы строения. ТИП РИЗОПОДЫ (RIZOPODA) В течение всего или большей части жизненного цикла органоиды движения — псевдоподии. Снаружи тело ее покрыто тонкой цитоплазматической мембраной (7,5 нм), за которой идет слой прозрачной довольно плотной эктоплазмы. Далее располагается зернистая полужидкая эндоплазма, составляющая основную массу тела животного. При формировании псевдоподии появляется сначала небольшой выступ эктоплазмы. Затем он растет и в него входит, как бы вливаясь, часть жидкой эндоплазмы. Псевдоподии амеб обычно не образуют разветвлении. Форма их может широко варьировал зависимости от условии среды (рH, температура, содержание солей в воде и др.). Разные виды амеб отличаются по числу и форме псевдоподий. Последние могут быть короткими, длинными, тупыми, заостренными на концах и т. п. Быстрота движения Amoeba proteus составляет при благоприятных условиях примерно 200 мкм в минуту. Передвигаясь, амебы наталкиваются на различные мелкие объекты: одноклеточные водоросли, клетки бактерий, частицы органического детрита, мелких простейших и т. п. Если объект достаточно мал амеба обтекает его со всех сторон, и он вместе с небольшим количеством жидкости оказывается внутри цитоплазмы простейшего. Таким образом, в эндоплазме образуются пищеварительные вакуоли. Внутрь последних из эндоплазмы поступают пищеварительные ферменты, и происходит внутриклеточное пищеварение. Жидкие продукты переваривания поступают в эндоплазму. Амебы обладают ферментами, расщепляющими белки, углеводы и липиды. Вакуоль с непереваренными остатками пищи подходит к поверхности тела и выбрасывается наружу. У амеб нет ни постоянного рта, ни порошицы 7 (отверстие для выведения экскрементов). У Amoeba proteus пищей служат другие мелкие простейшие и одноклеточные водоросли. Описанный способ заглатывания оформленной пищи при посредстве псевдоподий называется фагоцитозом. Наряду с ним недавно (главным образом благодаря применению для исследования электронного микроскопа) открыт другой способ поступления веществ снаружи в тело амебы, который называется пиноцитозом. Этим путем поглощаются жидкие вещества. Снаружи внутрь цитоплазмы впячивается тонкий канал, имеющий форму трубочки, в которую засасывается окружающая амебу жидкость. Затем образующаяся пиноцитозная вакуоль отшнуровывается и оказывается лежащей внутри цитоплазмы. При помощи этого механизма амеба как бы «пьет» жидкость. Обычно пиноцитозные каналы и вакуоли очень малы (меньше микрометра) и поэтому их удается рассмотреть лишь на ультратонких срезах с применением электронного микроскопа. Кроме пищеварительных вакуолей в теле амеб находится еще одна так называемая сократительная, или пульсирующая, вакуоль. Это пузырек водянистой жидкости, который периодически нарастает, а затем, достигнув определенного объема, лопается, опорожняя свое содержимое наружу. Вскоре на том же месте снова появляется маленькая капелька, проделывающая тот же цикл. Промежуток между двумя пульсациями вакуоли у разных амеб равен 1 — 5 мин. Основная функция сократительной вакуоли — регуляция осмотического давления внутри тела простейшего. Вода из окружающей среды проникает в тело амебы через наружную мембрану осмотически. Концентрация различных растворенных веществ в теле амебы выше, чем в пресной воде, благодаря чему создается разность осмотического давления внутри и вне тела простейшего. Однако в организме простейшего есть своего рода откачивающий аппарат, периодически выводящий избыток воды из тела, — сократительная вакуоль. Подтверждением этой функции вакуоли служит распространение их преимущественно у пресноводных простейших. У морских и паразитических форм, окруженных жидкостью с более высоким, чем в пресной воде, осмотическим давлением, сократительные вакуоли обычно отсутствуют или же сокращаются очень редко. Сократительная вакуоль кроме осморегуляторной функции частично выполняет и выделительную функцию, выводя вместе с водой в окружающую среду продукты обмена веществ. Однако основная функция выделения осуществляется непосредственно через наружную мембрану. Известную роль играет, вероятно, сократительная вакуоль и в процессе дыхания, ибо проникающая в результате осмоса в цитоплазму вода несет растворенный кислород. В эндоплазме амеб находится пузырьковидное ядро, богатое ядерным соком и обладающее всеми компонентами клеточного ядра — оболочкой, пронизанной многочисленными порами, ядерным соком, хроматином и одним или несколькими ядрышками. Существуют некоторые виды амеб, обладающие не одним, а несколькими ядрами. Многие виды свободноживущих амеб (особенно те, которые живут в почве) способны при наступлении неблагоприятных условий (например, 8 подсыхании) инцистироваться. При этом амеба втягивает псевдоподии, округляется и выделяет вокруг себя прочную двойную оболочку белковой природы. В таком неактивном состоянии амебы могут сохраняться долгое время (месяцами), а затем вновь переходить к активной жизни. Размножение. Амебам свойственно бесполое размножение, осуществляемое путем деления надвое. Этот процесс начинается с митотического деления ядра. На теле появляется легкий перехват, постепенно врезающийся в тело простейшего и перешнуровывающий ее на две дочерние особи. В период деления прием пищи обычно прекращается. Кишечные амебы человека и их значение. В кишечнике человека и ряда позвоночных обитает большое количество видов паразитических амеб, которые питаются содержимым кишечника, бактериями и большей частью не причиняют никакого вреда хозяину. Примером может служить кишечная амеба человека — Еntаmоеbа соli. Однако среди обитающих в кишечнике человека амеб имеется один вид — дизентерийная амеба — Еntаmoebа histolytica, который может быть возбудителем тяжелой формы кишечного колита — амебиаза. Амеба эта имеет 20 — 30 мкм в диаметре, подвижна. Живет она в толстых кишках человека и обычно питается бактериями, не нанося никакого вреда. Подобное явление, когда патогенный паразитический организм не проявляет своей патогенности, называется носительством. Но в ряде случаев дизентерийная амеба начинает вести себя иначе: проникает под слизистую оболочку кишки, начинает там питаться и усиленно размножаться. Слизистая кишечника изъязвляется, результатом чего бывает тяжелый кровавый понос (колит). Распространение кишечных амеб осуществляется при помощи цист, выходящих вместе с фекальными массами наружу. Цисты очень стойки и длительное время сохраняют жизнеспособность и инвазионность (способность к заражению при попадании в кишечник человека). По строению цист можно установить вид амебы. Кишечная амеба Entamoeba coli имеет восьмиядерные цисты, тогда как дизентерийная (Entamoeba histolitica) — четырехъядерные. В цистах есть особые ярко окрашивающиеся включения — хроматоидные тельца. При сильном заражении с экскрементами выводится до 300 млн. цист в день. Кишечные амебы человека распространены по всему Земному шару. Подкласс Раковинные амебы (Testacealobosia). Представители этого отряда отличаются от амеб защитной раковиной, одевающей тело. Раковина Testacea обычно имеет вид округлого или овального мешочка с отверстием (устьем), из которого выдаются псевдоподии, имеющие у разных видов неодинаковую форму и длину. Раковина у одних форм представляет тонкий слой плотного органического вещества, у других она образуется посторонними частицами (песчинки и т. п.), склеенными выделениями цитоплазмы. Размножение раковинных амеб совершается, как и у представителей предыдущего отряда, делением надвое, причем одна из половин выходит наружу через устье и окружается новой раковиной. Раковинные корненожки распространены в пресных водах, встречаются преимущественно среди прибрежной растительности, на дне вблизи берега. 9 Значительное число видов Testacea живет в торфяных болотах. Тип Фораминиферы (Foraminifera). Строение и физиология. Фораминиферы, обитатели моря, устроены сложнее прочих корненожек. Число современных видов превышает 1000. Раковина фораминифер обнаруживает ряд этапов постепенного усложнения. В наиболее простых случаях раковина состоит из плотного органического вещества — псевдохитина. Это вещество выделяется эктоплазмой. У других видов к этой тонкой пленке приклеиваются захваченные псевдоподиями посторонние частицы главным образом песчинки. Получается хитиноидная основа, инкрустированная зернами кварца. Раковинки подобного типа массивны и тяжеловесны. У большинства современных фораминифер раковина тоже тонкой хитиноидной основы, но пропитанная углекислым кальцием. Обладая большой прочностью, раковины такого рода отличаются гораздо большей легкостью, чем инкрустированные. Форма раковины фораминифер чрезвычайно разнообразна. У некоторых видов раковина имеет форму продолговатого мешка, у других вытягивается в трубку, у третьих эта трубка закручивается в спираль. Все это однокамерные формы. Но у большинства фораминифер полость раковины поделена поперечными перегородками на камеры (многокамерные формы), которые сообщаются друг с другом отверстиями, имеющимися в перегородках. Взаимное расположение камер может быть различным. Они могут располагаться в один ряд, в два ряда, спирально и т. п. Каждая многокамерная корненожка начинает свою жизнь, будучи однокамерной, причем эта первая камера меньше позднейших и называется зародышевой. Отверстие, сообщающее раковину с внешним миром и служащее для выхода псевдоподий, называется устьем. Помимо устья у многих корненожек все стенки раковины пронизаны тончайшими порами, тоже служащими для выхода ложноножек. Строение псевдоподий у фораминифер, которые называются ризоподиями, чрезвычайно своеобразно. Они представляют собой длинные тонкие переплетающиеся и сливающиеся нити, образующие вокруг раковинки сложную сеть. В ризоподиях осуществляется непрерывный ток цитоплазмы. По одной и той же ризоподии одни струи ее текут в центростремительном (к раковине), другие — в центробежном направлениях. Ризоподии служат для улавливания и, частично, переваривания пищи, а также для передвижения животного. Они способны сокращаться и вытягиваться. Размножение. Чередование поколений. Фораминиферы обладают сложным жизненным циклом, в который включаются две формы размножения — бесполое и половое. Некоторые стороны жизненного цикла их были изучены лишь за последнее время. Рассмотрим в качестве примера цикл развития однокамерной корненожки Myxotheca arenilega. Описание цикла начнем с одноядерной стадии, которая называется гамонтом. Это название связано с ее дальнейшей судьбой. После некоторого периода свободной активной жизни ядро ее начинает многократно делиться, и корненожка становится многоядерной. Вокруг каждого из ядер обособляется небольшой участок цитоплазмы, и все тело корненожки распадается на множество мелких клеток, 10 каждая из которых формирует два жгутика неравной длины. Клетки выходят из раковины наружу в морскую воду и попарно сливаются. Таким образом, это половые клетки (гаметы), в результате слияния которых образуется зигота. Она дает начало новому поколению, формирующему раковину и называемую агамонтом. В агамонте происходит постоянное увеличение числа ядер в результате их митозов и сам он увеличивается в размерах. Зигота обладает диплоидным комплексом хромосом, так же как и развивающийся из зиготы агамонт. После завершения его роста происходит еще два деления ядер, которые являются редукционными (мейоз), и образующиеся в результате их ядра гаплоидны. Далее вокруг каждого ядра вновь обособляется участок цитоплазмы и все тело распадается на большое количество мелких одноядерных зародышей, именуемых агаметами (частица «а» по-гречески означает отрицание, так что их можно было бы назвать «негаметами»). Это бесполое размножение, которое ведет к образованию нового поколения с гаплоидным ядром. Каждая агамета окружается раковиной и дает начало гамонту — поколению, с которого мы начали рассмотрение цикла и который далее вновь образует гаметы. Таким образом, в жизненном цикле фораминиферы чередуются две формы размножения: половое (при посредстве гамет) и бесполое (через агаметы) и два поколения: гамонты (размножаются половым путем) и агамонты (размножаются бесполым путем). Особенно замечательно в жизненном цикле фораминифер относительно недавно открытое чередование гаплоидной и диплоидной фаз. Мы видели, что редукция (мейоз) происходит здесь не перед образованием гамет, как это бывает у всех многоклеточных животных, а при образовании стадий бесполого размножения — агамет. Это единственный случай в животном мире, когда одно поколение гаплоидно, а другое диплоидно. Напротив, в растительном мире, где закономерно чередуются спорофит и гаметофит, происходит также и чередование гаплоидной (гаметофит) и диплоидной (спорофит) фаз ядра. Разумеется, это сходство не говорит о связи фораминифер с растениями, но представляет интересный пример конвергентного развития. В деталях (строение гамет, их число, судьба гамонтов и т. п.) жизненные циклы фораминифер очень разнообразны, и мы не можем останавливаться на их подробном рассмотрении. Однако у всех изученных видов он слагается из двух поколений — полового и бесполого. Большинство Foraminifera живет на дне водоемов, иногда на глубинах в тысячи метров, питаясь разными мелкими организмами. Лишь немногие виды, например Globigertna, входят в состав планктона. Раковинки этих видов снабжены обычно длинными радиальными шипами, сильно увеличивающими поверхность и позволяющими «парить» в толще воды. Палеонтология и практическое значение. Корненожки входят в состав очень многих морских отложений, начиная с древнейших кембрийских. Толстые слои известняков, мела, зеленого песчаника и некоторых других осадочных пород состоят преимущественно из раковин фораминифер. Благодаря малым размерам и большой распространенности корненожки во многих отношениях удобнее, чем остатки крупных организмов, для 11 определения возраста геологических пластов при поисках нефти и других полезных ископаемых. Каждый пласт имеет свою характерную фауну корненожек. Поэтому в каждом нефтеносном районе достаточно один раз установить залегание нефти в определенном пласте для того, чтобы и в других местах узнавать о вероятной близости нефти по исследованию корненожек в породе. Литература Шарова И.Х. Зоология беспозвоночных. Стр. 3-40, 54-60.