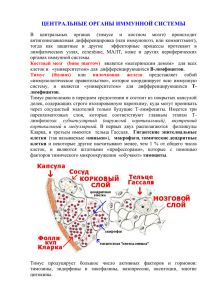
Разделение паренхимы на корковую и мозговую зоны
реклама

ТИМУС В ФИЛОГЕНЕЗЕ Селезнев Сергей Борисович Доктор ветеринарных наук, профессор Кузьменко Лариса Григорьевна Доктор медицинских наук, профессор 2011 Пути эволюции различных структур современных представителей животного мира и их преобразование в филогенезе можно проследить с помощью эволюционной морфоло-гии в сочетании с системным и историческим подходом. Исследования эволюции структур лимфоидной системы до конца ХХ века носили фрагментарный, а порой и противоречивый характер. ЭВОЛЮЦИЯ СТРУКТУР ИММУНОЛОГИЧЕСКОЙ ЗАЩИТЫ Б Е С П О З В О Н О Ч Н Ы Е Беспозвоночные Структуры иммунологической защиты Губки (2,5 млрд лет назад) Блуждающие амебоцициты Кишечнополостны (1 млрд лет назад) Амебоциты-макрофаги Неметрины (1 млрд лет назад) Лимфоцитоподобные амебоциты Кольчатые черви (1 млрд лет назад) Лейкопоэтические органы (узелки в целоме) Членистоногие (ракообразные) Димфомиелоидные образования в виде скопления компактных узелков вблизи глазной артерии Головоногие (моллюски) Лимфоэпителиальная железа («белое тельце»), напоминающая тимус у прзвоночных Таким образом, у беспозвоночных с полостью тела иммунная система обогатилась важным эволюционным элементом – «белым тельцем». В этом образовании происходят процессы ЛИМФО-МИЕЛОПОЭЗА. По своему строению «белое тельце» напоминает органы иммунной системы позвоночных. Снаружи оно покрыто соединительнотканной оболочкой, под которой располагается мозговое вещество. Последнее представлено лимфоидными тяжами и синусоидами. Основу лимфоидных тяжей образует рыхлая соединительная ткань. ЭВОЛЮЦИЯ ПОЗВОНОЧНЫХ ТИМУС У ПРЕДСТАВИТЕЛЕЙ ВОДНОГО МИРА Представители позвоночных Миксины Тимус Общая характеристика Не сформирован Расположение – Миноги Примитивный Дорсальная часть жаберных карманов Хрящевые рыбы Различаются корковая и мозговая зоны В головной части вблизи глазных впадин Костные рыбы Связан с эпителием глоточных кар-манов в Под жабрами ЭВОЛЮЦИЯ ТИМУСА У ПРЕДСТАВИТЕЛЕЙ ВОДНОГО МИРА Представители позвоночных Структурная организация тимуса Разделение на строму и паренхиму Разделение паренхимы на корковую и мозговую зоны Присутствие тимических телец (телец Гассаля) Присутствие не тимических клеточных элементов Миноги Нет Нет Нет Нет Хрящевые Имеется рыбы Имеется У некоторых представителей имеются скопления клеток Костные рыбы Имеется Имеется (за исключением лососевых) Встречаются ? Тучные клетки; в капсуле и септах – гранулоциты, макрофаги Тимус у земноводных: безногих, хвостатых и бесхвостых амфибий Тимус у бесхвостых амфибий располагается на боковой стороне черепа позади барабанной перепонки; состоит из 3 долей; формируется из эпителия второго глоточного кармана; корковая и мозговая зоны разделены клеточным барьером; в клеточном барьере располагается большое количество плазматических клеток и кровеносных сосудов; в корковой зоне находятся пролиферирующие лимфоциты. У этого отряда позвоночных Т-лимфоциты уже дифференцируются в тимусе, а В-лимфоциты – в костном мозге. Тимус у рептилий Тимус у черепах расположен в вентральной части шеи, состоит из отдельных долек; у ящериц, как и у змей, расположен анало-гично, состоит из двух долей, разделенных на кору и мозговое вещество; тимус крокодила состоит из отдельных эллипсовидных долек, достигающих области сердца, в каждой из них имеется корковое и мозговое вещество; в коре выявляется большое количество лимфоцитов, окружен-ных звездчатыми эпителиальными клетка-ми; в мозговой зоне обнаруживаются лим-фоциты, макрофаги. Тимические тельца в в мозговой зоне рептилий не обнаруживаются. Время появления рыб, амфибий, рептилия в палеонтологической летописи прямо пропорционально структурной организации тимуса. Так, у ящериц (конец пермской системы) тимус, еще не разделен на дольки, в клоаке у них отсутствуют лимфоидные элементы; у крокодилов (конец триасовой системы) тимус четко разделен на дольки, и у них появляется зачаток клоакальной сумки Дивергенция млекопитающих и птиц Рептилии Млекопитающие Птицы Птицы Тимус птиц располагается в облас-ти шеи и состоит из двух долей, каждая из которых делится на не-сколько овальных долек серо-розо-вого цвета. В тимусе птиц различают субкапсу-лярную, кортикальную и медулляр-ную зоны. В медуллярной зоне выявляются тимические тельца. Клоакальная сумка птиц располагается в грудо-брюшной полости под позвоночным столбом. У птиц происходит дальнейшее совершенствование иммунной системы. У них (и только у них !) появляется клоакальная сумка, полностью контролирующая процессы пролиферации, дифференцировки и созревания Влимфоцитов. Клоакальная (Фабрициева) сумка у птиц представляет собой вершину эволюционной диффе-ренцировка, поскольку у них имеет место топо-графическое и органное обособление части си-стемы, ответственнной за дифференцировку В-лимфоцитов (А.А. Заварзин, 1985) Млекопитающие Млекопитающие, как и птицы, произошли от рептилий, но линия, ведущая к млекопитающим дивергировала от родословного дерева в триасовый период (ок.250 млн лет назад). Система иммунной защиты млекопитающих стоит ближе к рептилиям, чем к птицам. Центральными органами иммунной системы у млекопитающих, как и у рептилий, являются тимус и костный мозг. Тимус млекопитающих, как и у птиц, состоит из двух долей, форма которых значительно варьирует. У парнокопытных он имеет лентовидную форму, у хищников – конусовидную, у приматов – пирамидальную. В тимусе у млекопитающих различают тело и 4 рога: 2 шей-ных остроконечных и 2 грудных, образующих основание тимуса. Паренхима тимуса дифференцируется на 4 зоны: субкапсу-лярную, кортикальную, медуллярную и внутридольковые периваскулярные пространства. Все зоны имеют различное функциональное значение. В целом можно отметить, что сравнительный анализ органных структур иммунной системы демонстрирует историческую направленность процесса по линии морфологического прогресса. Особое внимание обращают 3 события: Возникновение амебоцита-макрофага (и на его основе – лимфоцита (как морфологически обособленного клеточного типа); формирование лимфомиелоидного комплекса как самостоятельного образования; разделение лимфоидного комплекса и выделение лимфоидных структур для формирования самостоятельных, отдельных от кроветворения образований. - - Становление специфических иммунных реакций в филогенезе Система биологической защиты выс-ших животных и человека основана на функционировании многокомпо-нентной системы, включающей как неспецифические факторы защиты, так и специфические иммунные механизмы БЕСПОЗВОНОЧНЫЕ Главные механизмы биологической защиты – фагоцитоз и целомическая жидкость. Морфологический состав клеток, ответственных за фагоцитоз: амебовидные клетки крови и целомичес-кой жидкости (ацидофильные и базофильные клетки, бактериолизины, гемолизины, агглютинины). Кораллы и кольчатые черви способны распознавать и отторгать трансплантаты. Синтез АНТИТЕЛ у беспозвоночных отсутствует. Миксины и миноги У МИКСИН появляется способность распознавать аллогенные лимфоциты в смешанной культуре благодаря MДС-реакции (аналог реакции «трансплан-тат против хозяина»), которая характеризует дотими-ческий уровень развития клеточного иммунитета. МОРСКАЯ МИНОГА отличается наличием в крови лимфоцитов, сходных с малыми лимфоцитами высших животных, которые способны отвечать на ограниченное количество чужеродных антиге-нов, но никакая стимуляция не приводит к появлению у них плазматических клеток РЫБЫ Лимфоциты ХРЯЩЕВЫХ РЫБ под воздействием Con A и ФГА способны подвергаться бластной трансформации. У КОСТНЫХ РЫБ в селезенке, головной почке и тимусе выявляются плазматические клетки, синтезирующие антитела; у двоякодышащих рыб впервые появляется IgN (120000 D) – способен фиксировать комплемент, подобно IgM высших животных; у камбалы впервые в филогенетическом ряду появляется гиперчувствительность немедленно-го типа. ЗЕМНОВОДНЫЕ У АМФИБИЙ количество антителообразование зависит от температуры окружающей среды, но количество антителообразующих клеток от тем-пературы зависимости не имеет. У этих представителей появляются плазматичес-кие клетки, связанные с кишечником и похожие на плазматические клетки млекопитающих. Низшие амфибии (саламандры) кожные транс-плантаты отторгают медленно, бесхвостые (лягушки) – быстро; жабы способны развивать РБТЛ в смешанной культуре клеток; клетки японского тритона компетентны в осуществлении реакции «трансплантат против хозяина». Тимэктомия личинок амфибий приводит к значительному подавлению иммунных реакций ЗЕМНОВОДНЫЕ Большой интерес возникает о происхож-дении у амфибий Т- лимфоцитов. В экспериментальных работах показано и подтверждено с помощью хромосомного анализа, что первичная популяция Т-лимфоцитов проис-ходит из эпителиального зачатка тимуса, и этот орган у бесхвостых амфибий можно рассматривать как мощный источник предшествен-ников Т-лимфоцитов, по крайней мере на ранних этапах онтогенеза. АМФИБИИ И ПРОТИВООПУХОЛЕВЫЙ ИММУНИТЕТ Этот класс позвоночных способен вырабатывать противоопухолевый иммунитет. Известно, что у амфибий могут развивать-ся злокачественные опухоли. Полагают, что в процессе опухолеобразования важ-ную роль играет тимус. Так, устойчивые к опухоли аксолотли, становятся к ней чув-ствительными, если на ранних Рептилии Сведения о состоянии иммунной системы рептилий немногочисленны. Решающую роль в антителообразовании у этих представи-телей играет температура внешней среды. Она, в частнос-ти, имеет большое влияние на реакцию отторжения транс-плантата и реакцию «трансплантат против хозяина. Репти-лии, в отличие от амфибий, кожный трансплантат отторга-ют длительно. Рептилии имеют иммунологически отличные цепи для двух разных иммуноглобулинов, подобно IgM и IgG млекопитаю-щих, и в тот же время у них определяется IgN. У рептилий, в частности, у черепах при электронномикро-скопическом исследовании выявляются отличия от анало-гичных клеток млекопитающих. К числу таких отличий от-носится, например,то, что секреция иммуноглобулинов у них не имеет связи с обратным пиноцитозом и сопровож-дается разрушением ПТИЦЫ и МЛЕКОПИТАЮЩИЕ Основные особенности иммунной системы у представите-лей данных классов: 1) выражена функциональная гетерогенность лимфоци-тов; 2) иммунный ответ к большинству антигенов осуществля-ется с помощью кооперации Т-, Влимфоцитов и макрофа-гов; 3) в ходе иммунного ответа вырабатываются высокоаффинные и высокоспецифичные антитела, принадлежащие к классам A, M, G; 4) у млекопитающих, помимо этого, синтезируются анти-тела классов E и D. Для высших животных и человека в настоящее время общепризнанным является следующее: 1) родоначальником всех клеток иммунной системы является полипотентная стволовая кроветворная клетка; 2) она не выполняет никаких специализированных функций, но может делиться неограниченное количество раз; 3) одна из дочерних клеток остается стволовой, другая диф-ференцируется; 4) стволовые клетки, поступающие в тимус, дифференциру-ются в Т-лимфоциты. Несмотря на то, что трансплантационный иммунитет и кооперация клеточных элементов впервые возникли у беспозвоночных, то с появлением специализированного органа, ответственного за обеспечение клеточного иммунитета, усложнением его структуры и увеличением степени функциональной активности, иммунная система млекопитающих (в том числе и человека) наибольшей степени совершенства. Благодарю за внимание!