и “закрытых” системах мозга
реклама

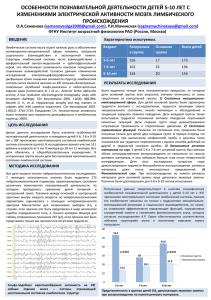
Вестник РАУ. Серия физико-математические и естественные науки, 2, 2006, 103-108 103 УДК 612. 826 СЕНСОРНАЯ ИНФОРМАЦИЯ В “ОТКРЫТЫХ” И “ЗАКРЫТЫХ” СИСТЕМАХ МОЗГА З.А. Ваграмян Институт прикладных проблем физики НАН РА Современная нейрофизиология, направляющаяся в сторону раскрытия процессов образования и передачи информации, использует и развивает системный подход в оценке явлений. Собственный систематически исследованный экспериментальный материал по изучению электрической активности лимбической коры кошки и взаимодействия разных структур лимбической системы, а также литературные сведения привели нас к представлению о наличии систем мозга, отличающихся по рецептивно-информативной способности, и представленных условно в виде “открытых” и “закрытых” систем мозга, в которых проведение сенсорной информации и ее обработка происходят по-разному. В отличие от “открытых” систем мозга, несущих одномодальные сигналы, “закрытые” системы при наличии многоканальных возможностей могут конвергировать на одном нейроне разномодальные сигналы и, следовательно, нести функцию наивысших интеграторов мозга. Опираясь на большие успехи, достигнутые методами электрического стимулирования нервной ткани и регистрации протекающих в ней электрических процессов, нейрофизиологи стремятся раскрыть и понять механизмы деятельности мозга. В поисках нового современная нейрофизиология, по-видимому, направляется в сторону раскрытия процессов образования и передачи информации, лежащих в основе интегративных механизмов управления нервными процессами. Известный нейрофизиолог Шеперд в своей книге “Нейробиология” пишет: “В настоящее время появились новые способы описания корковых областей: мы подразделяем их по организации внутренних сетей, по входным и выходным связям, по специфическим функциональным свойствам и нейрохимическим особенностям. Каждое поле отличается от соседних полей по функциональным свойствам и по связям в пределах определенной системы” [1]. В то же время ученые мира считают, что интегративные возможности нейронных сетей все еще лежат за пределами наших представ- 104 Сенсорная информация в “открытых” и “закрытых” системах мозга лений, однако каждый научный кирпичик, вставленный в эту монументальную проблему, является ценным вкладом в ее развитие. Для того, чтобы приблизиться к рассмотрению интегративных механизмов деятельности разных систем мозга, необходимо представить некоторые взаимоотношения внутримозговых систем, принимающих разнообразный поток информации. Понятие “системы” представляется современному нейрофизиологу как многоуровневый ансамбль взаимосвязанных структур, ответственных за реализацию определенной функции. Систему составляют “нервные сети”, объединяющие популяции функционально взаимосвязанных нейронов. Сенсорные сети включаются в деятельность при поступлении внешнего сигнала в соответствующее рецепторное поле. Согласно нашему мнению, внутримозговые системы по рецептивно-информативной способности можно условно разделить на две группы: “открытые” и “закрытые” системы. К числу “открытых” систем мозга можно отнести те, которые пользуются периферическим входом или рецептивным полем, непосредственно контактирующим с внешней средой. К “открытым” системам мозга относятся филогенетически самые древние, наиважнейшие системы: обонятельная, вкусовая, осязательная, зрительная и слуховая. В настоящее время считается, что кроме “пяти чувств” мы чувствительны к таким модальностям как температура, боль и вибрация. Модальностью также является чувство равновесия и др. Поэтому бесспорно, число модальностей намного превосходит пять чувств, хотя их нельзя пока точно установить [1, 2]. Входы “открытых” систем пользуются натуральными сигналами, например, слуховая система воспринимает звуковые сигналы через рецепторы ушной раковины, несущие информацию о параметрах раздражителя и т. д. В то же время надо обратить внимание на то, что нерв передает специфическую модальность, независимо от того, что его стимулирует. К примеру, можно ощутить звук в ушах и в результате удара по голове, и при электрической стимуляции нерва. Таким образом, “открытые” системы мозга пользуются лишь только модально-специфическими сигналами, ибо рецепторы, как правило, специализированы и отвечают только на один тип раздражителя. Нервные сенсорные сети, а, следовательно, мозговые “открытые” системы функционируют благодаря поступлению сигналов в рецепторные клетки, составляющие рецептивные поля. Общая функциональная особенность всех рецепторных клеток состоит в том, что они превращают одну форму энергии в другую, например, чувствительные окончания кожи преобразуют механическую и тепловую энергию в электрическую энергию нервного потенциала [2, 3]. Рецепторные клетки кожи (и подкожной клетчатки) обеспечивают большой объем информации об особенностях предмета, с которым они приходят в соприкосновение. В “открытых” системах первичные рецепторные афференты вносят контролирующую коррекцию и влияют на поступление сенсорной информации. Сенсорные нервные окончания выполняют функцию преобразователя, трансформируя различные формы физической энергии в серию потенциалов З.А. Ваграмян 105 действий, возникающих в аксонах. Эти разряды представляют сенсорный код. Итак, рецепторы работают как биологические преобразователи, превращающие энергию раздражителя в электрическую энергию нервного импульса. В функции преобразователя, выполняемой рецепторной клеткой, при регистрации электрических реакций наблюдается наличие двух потенциалов: первый – градуальный ответ, и второй – генераторный потенциал действия, обеспечивающий “запуск” импульса в аксоне [3]. Итак, в рецепторах начинается процесс кодирования. “Закрытые” системы мозга как интеграторы разномодальных сигналов Те системы мозга, которые получают опосредованные сигналы извне, могут быть отнесены к системам “закрытого” типа. В частности, к внутримозговым системам “закрытого” типа относятся “эмоциональная” и система “памяти”. Системы “закрытого” типа работают в режиме сбора модально-неспецифических, уже переработанных на уровне рецепторов, сигналов из многих других систем, в том числе и из “открытых”, что свидетельствует об интегративной деятельности “закрытых” систем [4 – 6]. Системы “закрытого” типа пользуются ассоциативными связями, образующими ассоциативные поля мозга. Как известно, качество и значение нервного сигнала зависит от того, где берут начало и где оканчиваются нервные волокна, т.е. от их связей [7]. Переработка информации происходит на иерархически организованных уровнях, где сигналы переходят от одной функциональной группы клеток к другой. Поэтому функциональное значение какой-то группы нейронов можно выяснить, изучая эффекты, возникающие в результате конвергенции от снабжающих ее нервных волокон. Примером может послужить конвергентная способность лимбической коры [8, 9], относящейся к типу “закрытых” систем. На рис. 1 представлены постстимуляторные гистограммы, отражающие взаимодействие трех структур лимбической системы, в частности, влияние стимуляции амигдалы (рис. 1, 1) и гипоталамуса (рис. 1, 3) на электрическую активность нейрона лимбической коры (поле 24), учитывая, что включение данной системы происходит именно в ответ на стимуляцию лимбических структур. Анализ постстимуляторных гистограмм ответов фоновоактивных нейронов лимбической коры на раздражение задне-латерального гипоталамуса и центрального ядра амигдалы выявил высокий уровень конвергенции импульсов 70,7% (70 нейронов из 99). Обнаружение бимодальных нейронов лимбической коры, отвечающих на раздражение гипоталамуса и амигдалы, по-видимому, показывает, что в некоторых случаях к одному и тому же нейрону лимбической коры подходят качественно различные волокна и окончания от разных структур. Таким образом, нами экспериментально показано, что в лимбическую кору (верхний этаж эмоционального мозга) направляются конвергирующиеся на одном и том же нейроне возбудительные импульсы в ре- 106 Сенсорная информация в “открытых” и “закрытых” системах мозга зультате стимуляции гипоталамуса и амигдалы, относящихся к мотивационной системе мозга [10]. Рис. 1. Ответы нейрона лимбической коры на стимуляцию задне-латерального гипоталамуса и центрального ядра амигдалы. 1–4, постстимуляторные гистограммы распределения плотности потенциалов. 1 – фоновая активность нейрона (поле 24), 2 – при стимуляции заднего гипоталамуса, 3 – фоновая активность нейрона, 4 – при стимуляции центрального ядра амигдалы. Ширина бина – 2 м/сек., 10 реализаций, калибровочные сигналы – 5 имп. 50 м/сек. Система в системе Г. Шеперд указывает на существование микросистем в системах. Это понятно. В то же время можно обратить внимание на то, что одним из основных компонентов “эмоциональной” (“закрытой”) системы является чувствительная (“открытая”) система. Пути разных модальностей (болевой, температурной и тактильной) чувствительности начинаются на уровне рецепторных образований на поверхности тела. Сенсорные сигналы этих видов чувствительности возникают и передаются по нервным волокнам клеток спино-мозговых узлов, центральные отростки которых образуют задние корешки спинного мозга. В задних рогах спинного мозга начинается важный восходящий путь, который достигает соответствующих областей коры головного мозга, переключаясь в ядрах таламуса, относящихся к филогенетически старой части зрительного бугра (палеоталамус), например, к центральному латеральному ядру таламуса, которое посылает волокна в лимбическую кору (поясную извилину). Так, чувствительные волокна достигают лимбической коры, таким образом принимая участие в реализации механизмов эмоциональной системы мозга. Волокна висцеральной болевой чувствительности образуют такие же связи в задних рогах спинного мозга, как и волокна соматической болевой чувствительности. Однако волокна, проводящие висцеральную болевую чув- З.А. Ваграмян 107 ствительность, образуют более диффузные системы. Вероятные причины этого явления Шаде и Форд приводят в своих интерпретациях. В заключение необходимо отметить, что передача сенсорной информации в “открытых” и “закрытых” системах мозга происходит благодаря наличию чрезмерно большого количества сигналов, т.е. их избыточности. Согласно теории информации, чем больше избыточность при кодировании, тем надежнее застрахована от искажений передаваемая информация. К тому же, параллельная передача информации по двум или большему числу каналов рассматривается как использование избыточности и защита от шума. Из-за дивергенции и конвергенции на переключательных синапсах параллельные каналы могут оказаться поперечно соединенными, за счет чего избыточность возрастает. Таким образом, приведенные выше данные, как нам кажется, позволяют представить наличие относительно “открытых” и “закрытых” систем мозга, отличающихся по своей рецептивно-информативной способности, и, следовательно, с различной функциональной специализацией в мозгу. В отличие от “открытых” систем, несущих одномодальные сигналы, “закрыты” системы при наличии многоканальных возможностей могут конвергировать на одном нейроне разномодальные сигналы и, следовательно, нести функцию наивысших интеграторов мозга. ЛИТЕРАТУРА 1. 2. 3. 4. G. Shepherd Нейробиология (русск. перев.).- М.: Мир, 1987, с. 450. J.P. Schade, D.H. Ford, Basic Neurology, London, New York. 1976, p. 344. H. Tamar, Основы сенсорной физиологии (русск. перевод), 1976, с. 510. Z.A.Vagramyan, O.G. Baklavadjian, Activity of Neutrons in Limbic Cortex during Stimulation of Somatosensory Zones. // J. Neuroscience and Behavioral Physiology, 1986, v. 16. N 1, p. 23-28. 5. З.А. Ваграмян, Особенности нейронной организации амигдало-цингулярных взаимоотношений. // Вестник МАНЭБ, 2001, вып. 2, N 6, с. 140-142. 6. З.А. Ваграмян, Организация и регуляция электрической активности лимбической коры. // Информационные технологии и управление. 2002, N 4-1, с. 10-13. 7. S.W. Kuffler, J.G. Nicholls, From neutron to brain. 1979, с. 430. 8. З.А. Ваграмян, Сравнительный анализ влияний амигдалы и гипоталамуса на активность нейронов лимбической коры. / Материалы докладов VI съезда арм. физиологов общества им. Л.А. Орбели, Ер., 2001, с. 90-93. 9. Z.A.Vahramyan, Neuronal organization of the limbic cortex. / Third conference of the Arm. IBRO Association. Yerevan 2000, p. 75-76. 10. П.В. Симонов, Лекции о работе головного мозга.- М.: Наука, 2001, 100 с. Поступила в редакцию 16. 06. 2006 108 Сенсорная информация в “открытых” и “закрытых” системах мозга ¶ÈÊàôÔºÔÆ §´²ò¦ êÆêîºØܺðÆ ¼¶²Ú²Î²Ü ÆÜüàðزòÆ²Ü ¼.². ì³Ññ³ÙÛ³Ý Ðá¹í³ÍáõÙ ÷áñÓÝ³Ï³Ý ¨ ·ñ³Ï³ÝáõÃÛáõÝÇó ѳÛïÝÇ ïíÛ³ÉÝ»ñÇ ÑÇÙ³Ý íñ³ Ý»ñϳ۳óíáõÙ ¿ ï»ë³Ï»ï, Áëï áñÇ ³Ûëå»ë Ïáãí³Í §μ³ó¦ ¨ §÷³Ï¦ áõÕ»ÕÇ Ñ³Ù³Ï³ñ·»ñÁ ï³ñμ»ñíáõÙ »Ý Áëï Çñ»Ýó §ÁÝϳÉÙ³Ý (é»ó»åïÇí) ¨ ÇÝýáñÙ³óÇáݦ ѳïÏáõÃÛáõÝÝ»ñÇ: Æ ï³ñμ»ñáõÃÛáõÝ §μ³ó¦ ѳٳϳñ·»ñÇ, áñáÝù û·ïíáõÙ »Ý Ù»Ï ï»ë³ÏÇ ³½¹³Ý߳ݳÛÇÝ Ñ³Ù³Ï³ñ·Çó, §÷³Ï¦ ѳٳϳñ·»ñÁ û·ï³·áñÍáõÙ »Ý μ³½Ù³³½¹³Ý߳ݳÛÇÝ Ñݳñ³íáñáõÃÛáõÝÝ»ñ ¨ ³Û¹åÇëáí ϳï³ñáõÙ »Ý áõÕ»ÕÇ μ³ñÓñ³·áõÛÝ Ñ³í³ù³Ï³Ý, ÇÝï»·ñ³ïÇí ýáõÝÏódzÝ: THE SENSORY INFORMATION IN “OPEN” AND “CLOSED” BRAIN SYSTEMS Z.A. Vahramyan The presence of “open” and “closed” brain systems is considered in this article. These two types of systems are distinguished by their receptive-informative ability. Unlike on “open” brain systems, which conduct only monomodal signals, “closed” systems converge polymodal signals on the same neutron, that is perform the brain higher integrators’ function.