На правах рукописи Специальность 03.03.04 Клеточная биология, цитология, гистология
реклама

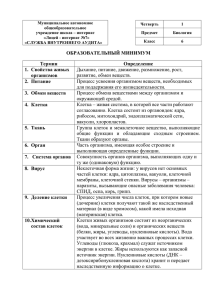
На правах рукописи Гаранина Анастасия Сергеевна ЭНТОЗ В СУБСТРАТ-ЗАВИСИМЫХ КУЛЬТУРАХ НОРМАЛЬНЫХ И ОПУХОЛЕВЫХ КЛЕТОК ЧЕЛОВЕКА Специальность 03.03.04 Клеточная биология, цитология, гистология АВТОРЕФЕРАТ диссертации на соискание ученой степени кандидата биологических наук Москва – 2015 Работа выполнена на кафедре клеточной биологии и гистологии биологического факультета Федерального государственного бюджетного образовательного учреждения высшего образования «Московский государственный университет имени М.В. Ломоносова». Научный руководитель: Онищенко Галина Евгеньевна, доктор биологических наук, профессор, заведующий кафедрой клеточной биологии и гистологии биологического факультета Федерального государственного бюджетного образовательного учреждения высшего образования «Московский государственный университет имени М.В. Ломоносова». Официальные оппоненты: Александрова Антонина Юрьевна, доктор биологических наук, ведущий научный сотрудник лаборатории механизмов канцерогенеза Федерального государственного бюджетного учреждения «Российский онкологический научный центр имени Н.Н. Блохина» Министерства здравоохранения РФ, Москва. Андреева Елена Рамуальдовна, кандидат биологических наук, ведущий научный сотрудник лаборатории клеточной физиологии Федерального государственного бюджетного учреждения науки «Государственный научный центр Российской Федерации Институт медико-биологических проблем» РАН, Москва. Ведущая организация: Федеральное государственное бюджетное учреждение науки «Институт биологии развития им. Н.К. Кольцова РАН». Защита состоится 15 марта 2016 года в 15.30 на заседании диссертационного совета Д 501.001.52 при Московском государственном университете имени М.В. Ломоносова по адресу: 119234, Москва, Ленинские Горы, д.1, стр.12, Биологический факультет МГУ, аудитория М-1. С диссертацией можно ознакомиться в Научной библиотеке Московского государственного университета имени М.В. Ломоносова и на официальном сайте биологического факультета МГУ http://www.bio.msu.ru. Автореферат разослан 2016 г. Ученый секретарь диссертационного совета Калистратова Е.Н. 2 ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ Актуальность темы исследования. Программированная гибель клеток (ПКГ) – неотъемлемая часть жизни многоклеточных организмов. В 2009 г. в список разновидностей ПКГ был впервые включён энтоз – вариант клеточного каннибализма (КК), при котором клетка внедряется внутрь другой клетки и там погибает. До сих пор процессы, лежащие в его основе, детально не изучены. Имеющиеся данные получены, как правило, на опухолевых клетках, переведённых в суспензионное состояние. Существуют различные мнения о роли КК. Он может способствовать увеличению плоидности клеток и выживанию клеток злокачественной опухоли при неблагоприятных условиях среды. КК также позволяет опухолевым клеткам уходить от иммунного ответа организма. Всё это способствует росту и малигнизации опухоли. В связи с последним предполагается, что энтоз можно использовать для диагностики опухолевой прогрессии. Таким образом, исследование энтоза – это перспективное направление фундаментальной и прикладной биологии. В первом случае изучение данного типа КК позволит получить более полную и глубокую информацию о процессах, осуществляющих этот вариант ПКГ, что расширит представление о путях клеточной смерти. С точки зрения прикладной науки выявление процессов, включённых в реализацию энтоза, предоставит возможность управлять этим явлением, что может быть использовано для терапии злокачественных новообразований, а также откроет новый способ диагностики опухолей. Степень разработанности темы. Описано два вида энтоза – гомо- и гетеротипический, при которых взаимодействуют клетки одного или разных типов, соответственно (Kroemer and Perfettini, 2014; Sun et al., 2014). Ключевую роль в обоих случаях играет акто-миозиновый комплекс внедряющейся клетки (Overholtzer et al., 2007). Для осуществления внедрения клетки в клетку необходима также динамическая реорганизация микротрубочек, регулирующая упругость клеток (Xia et al., 2014). Судьба же внедрившейся клетки – это, как правило, лизосомо-опосредованная деградация. Перечисленные выше данные в основном получены на клетках, экспериментально переведённых в суспензионное состояние. Цель и задачи исследования. В связи с выше сказанным цель работы состояла в исследовании энтоза в субстрат-зависимых культурах нормальных и опухолевых клеток человека. Были поставлены следующие задачи: 1. изучить состояние адгезивных контактов между внутренней и наружной клетками в ходе гомотипического энтоза, а также исследовать локализацию и функциональное состояние ряда мембранных органелл клеток и компонентов цитоскелета; 2. установить роль элементов цитоскелета и мембранных органелл внутренней и наружной клеток в осуществлении гомотипического энтоза; 3 3. определить способ деградации внедрившейся клетки при гомотипическом энтозе; 4. выявить возможность гетеротипического энтоза при сокультивировании нормальных и опухолевых клеток субстрат-зависимых культур; 5. установить, какие варианты энтозов – гомо- или гетеротипический – преобладают в этих условиях; 6. определить способ деградации внедрившейся клетки в ходе гетеротипического энтоза. Научная новизна. В работе впервые выявлен энтоз в человеческих культурах нормальных и опухолевых клеток эпидермального происхождения в прикреплённых к субстрату условиях. Получены новые данные о наличии десмосом между клеткамиучастницами энтоза. Установлено, что в процессе внедрения клетки в клетку важную роль играет актиновый цитоскелет как внедрившейся клетки, так и наружной. Получены оригинальные данные о роли микротрубочек и аппарата Гольджи при энтозе. На основе морфологических изменений клеток-участниц энтоза данный процесс разделён на 5 стадий после внедрения клетки в клетку, что позволит систематизировать описание исследуемого варианта ПКГ. Выявлена возможность гетеротипических взаимодействий между нормальными и опухолевыми клетками эпителиального происхождения и проведено сравнение частоты встречаемости различных вариантов энтоза при их сокультивировании. Предложен новый способ доказательства локализации одной клетки внутри другой. Составлена схема последовательности событий после внедрения клетки в клетку, связанных с особенностями строения энтозной вакуоли. Впервые разработан метод синхронизации вступления клеток в процесс энтоза. Теоретическая и практическая ценность работы. Результаты работы содержат новые сведения о поведении клеток, а также о функциях их органелл в ходе такого варианта ПКГ, как энтоз. Полученные данные могут быть использованы в исследованиях, направленных на изучение процессов ПКГ. Также они могут открыть новые возможности в разработке подходов противоопухолевой терапии. Методология и методы исследования. В работе применяли классические цитологические методы получения и анализа данных, такие как: культивирование эукариотических клеток, прижизненное окрашивание и наблюдение за клетками, фиксация, цито- и иммуноцитохимическое окрашивание, ингибиторный анализ, мечение клеток бромдезоксиуридином, метод конфокальной сканирующей лазерной микроскопии, методы электронной микроскопии, статистический анализ результатов. Кроме того, был использован метод корреляционной микроскопии. Личное участие автора. Работа полностью выполнена автором, включая анализ научной литературы, разработку экспериментальной части, получение и обработку результатов. 4 Степень достоверности и апробация результатов. Достоверность результатов данной диссертации подтверждается анализом современного состояния вопроса, адекватностью методических подходов, большим объёмом воспроизводимых результатов и статистическим анализом данных. Материалы работы представлены на 2 всероссийских и 4 международных конференциях: I всероссийская конференция «Внутриклеточная сигнализация, транспорт, цитоскелет» (Санкт-Петербург, 2011), VIII конференция молодых учёных ИБР РАН (Москва, 2012), XVIII и XXI международные научные конференции студентов, аспирантов и молодых учёных «Ломоносов» (Москва, 2011 и 2014), I международная научная конференция студентов и аспирантов Cell Technology Week (Киев, 2013), Международная научная конференция «Актуальные вопросы морфогенеза в норме и патологии» (Москва, 2014). По результатам диссертации опубликовано девять печатных работ, две из которых – статьи в рецензируемых журналах, соответствующих Перечню ВАК. Структура диссертации. Работа изложена на 165 листах машинописного текста и дополнена иллюстративным материалом в количестве 81-ого рисунка и 13 таблиц. Она состоит из введения, обзора литературы, описания материалов и методов, результатов, их обсуждения, заключения, выводов, списка сокращений и библиографического списка. ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ Материалы и методы Биологический материал. Исследования проводили на клетках линий А431 (эпидермоидная карцинома человека), MCF7 (аденокарцинома молочной железы человека) (Институт Цитологии РАН) и HaCaT (нормальные кератиноциты человека – минимально трансформированные клетки; любезно предоставлены Е.А. Воротеляк (Институт биологии развития им. Н.К. Кольцова РАН)). Клетки культивировали во флаконах или чашках Петри (SPL Lifesciences) в среде DMEM (ПанЭко) с добавлением 10% фетальной сыворотки телёнка (Австрия), 2 мМ L-глутамина (ПанЭко) и антибиотика-антимикотика (Sigma). В СО2инкубаторе (Sanyo) поддерживалась температура 37°С и 5% содержание углекислого газа. Морфологический анализ клеток. Клетки фиксировали раствором Буэна и окрашивали гематоксилином-эозином. Препараты исследовали на световом микроскопе Leica (объективы N Plan 100×/1,25, oil и 40х), оснащённом цифровой камерой DFC 295 и программным обеспечением Leica Аpplication Suite Version 3.4.0. Химические воздействия на клетки. Для разборки актиновых филаментов клетки обрабатывали цитохалазином В, затем отмывали культуральной средой 3 раза по 5 мин и продолжали культивировать. Для деполимеризации микротрубочек в среду культивирования добавляли нокодазол. При постановке эксперимента с брефелдином А клетки сначала 5 инкубировали с цитохалазином В в течение 48 часов (чтобы полностью ингибировать вступление в энтоз), затем отмывали и добавляли брефелдин А. Концентрация каждого из агентов в среде – 5 мкг/мл. В качестве контролей были взяты клетки, культивировавшиеся в обычной среде, либо в среде с диметилсульфоксидом (ПанЭко) в концентрации 5 мкг/мл. Было установлено, что ДМСО не оказывает влияния на исследуемые показатели. Прижизненные наблюдения за клетками. Для выявления лизосом использовали прижизненные красители акридиновый оранжевый (1 мкг/мл, Sigma) и LysoTracker Red DND99 (1:5000, Invitrogen); аутофагосом – моноданзилкадаверин (85,5 мкМ, Sigma); митохондрий – родамин 123 (1 мкг/мл, Sigma); АФК – 2’,7’-дихлорофлуоресцеиндиацетат (10 мкМ, BioChemika). Препараты исследовали на инвертированном флуоресцентном микроскопе Axiovert 200M («Carl Zeiss Inc.») (объектив PlanApo 63×/1,4, oil), снабжённом чёрно-белой цифровой камерой Carl Zeiss AxioCam и программным обеспечением AxioVision 3.1. Длительное прижизненное наблюдение проводили на линии MCF7 в матрасе с координатной сеткой на дне при помощи дифференциально-интерференционного микроскопа (DIC, Leica) (объектив HI Plan 20×/0,30) с цифровой камерой VideoZavr Standart 3.533 и программным обеспечением VideoZavr 1.4. Интервал между фотографиями – 1 ч. Мечение клеток бромдезоксиуридином. Для мечения клеток разных линий использовали BrdU (10 мкМ, Mannheim). Фиксация и цито-/иммуноцитохимическое окрашивание клеток. Клетки фиксировали 4% формалином. Пермеабилизация мембран осуществлялась с помощью Triton X100. При окрашивании клеток антителами к BrdU проводили дополнительную обработку 4N соляной кислотой. Для выявления полимеризованного F-актина использовали меченый TRITC фаллоидин (Sigma). Для иммуноцитохимического окрашивания – кроличьи антитела к βкатенину (Sigma, любезно предоставлены д.б.н. Кузнецовым С.А.), маннозидазе II (Abcam, любезно предоставлены Фокиным А.И.), активной каспазе-3 (Sigma), γ-тубулину (Sigma, любезно предоставлены д.б.н. Узбековым Р.Э.); мышиные антитела к плакоглобину (Sigma), белку аппарата Гольджи 58К (Sigma), α-тубулину (Sigma), BrdU (Sigma); овечьи антитела к цитохрому с (Sigma) и вторичные антитела, конъюгированные с FITC, TRITC, Alexa Fluor 488, Alexa Fluor 568 и Alexa Fluor 546. Ядра окрашивали DAPI (0,1 мкг/мл, Sigma). Препараты анализировали на микроскопе Axiovert 200M (объективы PlanApo 20× и 63×/1,4, oil), либо на конфокальном сканирующем лазерном микроскопе (Leica TCS SPE) (объектив ACS Apo 63x/1,3, oil) с программным обеспечением Leica Application Suite Version 2.8.1. 6 Сокультивирование клеток разных линий. Клетки одной из культур метили BrdU в течение 2-х суток. Затем клетки разных линий сливали в равных концентрациях и оставляли культивироваться в течение 1-х суток. Варианты сокультивирования были различны. Сканирующая и трансмиссионная электронная микроскопия (СЭМ и ТЭМ). Клетки фиксировали 2,5% глутаровым альдегидом с 2% формалином на 0,1М PBS, дофиксировали 1% OsO4 (для ТЭМ) и проводили по стандартной методике подготовки образцов для СЭМ и ТЭМ. Препараты исследовали на СЭМ SCAN JSM («JEOL»), ТЭМ JEM-1011 («JEOL») с цифровой фотокамерой GATAN ES500W, работающей под управлением программы Digital Micrograph фирмы GATAN, и пленочном ТЭМ JEM-100B («JEOL»). Корреляционная микроскопия. На фиксированном 2,5% глутаровым альдегидом с 2% формалином на 0,1М PBS и окрашенном DAPI препарате искали гомотипические энтозы с помощью фазово-контрастной микроскопии (Axiovert 200M) и отмечали места их локализации (при поиске гетеротипических энтозов проводили дофиксацию препаратов, окрашенных антителами к BrdU, 2,5% глутаровым альдегидом с 2% формалином на 0,1М PBS). Под DIC счищали слой клеток вокруг метки и проводили для СЭМ. Обработка и анализ результатов. Обработку фотографий проводили в программе а в ImageJ. Подсчёт числа митозов, апоптозов и энтозовб производили на 1000 клеток. Построение диаграмм и вычисление величин стандартного отклонения проводили в программе Microsoft Office Excel 2007. Статистическую обработку данных осуществляли в программе Statistica 6.0. Уровень значимости р вычисляли по U-критерию Манна-Уитни. 10 µm д г Результаты и обсуждение е Картины “клетка-в-клетке” в исследуемых культурах В субстрат-зависимых культурах А431, MCF7 и HaCaT были обнаружены картины гомотипического энтоза (Рисунок 1). ж б ав б 10 µm ! 10 µm 10 µm ! ги зв 10 µm л м к Рисунок 1. Гомотипический энтоз в культуре клеток НаСаТ. а – морфология клеток при энтозе, окрашивание гематоксилином-эозином, световая микроскопия; б-г – доказательство локализации клетки внутри другой клетки: б – фазово-контрастная микроскопия; в – цитохимическое окрашивание ядер с помощью DAPI, флуоресцентная 10 µm микроскопия; г – СЭМ. Зелёной стрелкой показана наружная клетка, красными – внутренние. ! 7 Внедрившиеся клетки (внутренние) имели округлую форму и находились внутри крупных неокрашиваемых энтозных вакуолей. В наружных клетках (энтозных) ядро приобретало бобовидную форму и смещалось к периферии клетки. Для доказательства локализации одной клетки внутри другой был использован метод корреляционной микроскопии (Рисунок 1 б-г). Частота встречаемости энтозов в культурах А431 и MCF7 составляла 1,5%. Для клеток линии НаСаТ энтотический индекс был значительно ниже – 0,5% на 1-е сутки культивирования. Процесс внедрения клетки в клетку На рисунке 2 представлены фотографии, предположительно иллюстрирующие различные этапы внедрения клетки в клетку. На начальном этапе одна из клеток (энтозная) прикреплена к субстрату, другая (внедряющаяся) – к первой клетке. Далее внедряющаяся клетка продавливает плазматическую мембрану энтозной клетки, в результате чего образуется впадина. По мере углубления данной впадины формируется кратер. На заключительном этапе внедрения энтозная клетка образует псевдоподию, которая накрывает внедряющуюся клетку. г в б a Рисунок 2. Внедрение клетки в клетку в субстрат-зависимой культуре А431. а – взаимодействие клеток; б – образование впадины; в – образование кратера; г – образование псевдоподии; СЭМ. Стадии энтоза в структурах “клетка-в-клетке” Чтобы систематизировать имеющиеся в литературе данные и сопоставить их с результатами нашей работы, мы разделили энтоз после внедрения клетки в клетку на 5 стадий на основе морфологических изменений внедрившейся клетки, обусловленных её деградацией (Рисунок 3). На первой стадии энтоза плазматическая мембрана внутренней клетки тесно контактирует с мембраной энтозной вакуоли. Ядро имеет округлую форму, богато диффузным хроматином, в нём чётко выявляются несколько ядрышек. На второй стадии на поверхности внутренней клетки обнаруживаются выросты в форме шипиков, чётко выявляются границы внутренней клетки и энтозной вакуоли. На третьей стадии очертания клетки становятся неровными, размер уменьшается, вследствие 8 чего расстояние между мембранами внедрившейся клетки и энтозной вакуоли значительно увеличивается. Ядро деформируется, хроматин начинает конденсироваться. На четвёртой стадии внутренняя клетка имеет неправильную форму, ядро сжато, хроматин заметно конденсирован, ядрышки не выявляются. На пятой стадии размер внутренней клетки значительно меньше исходного, пространство между плазматической мембраной внутренней клетки и мембраной энтозной вакуоли вновь сокращается (из-за уменьшения размеров энтозной вакуоли), ядро подвергается фрагментации. a 10 µm е б в г д ж з и к 10 µm Рисунок 3. Стадии энтоза в культуре клеток А431. а, е – первая стадия энтоза; б, ж – вторая стадия; в, з – третья стадия; г, и – четвёртая стадия; д, к – пятая стадия. а-д – фазовоконтрастная микроскопия; е-к – цитохимическое окрашивание ядер с помощью DAPI, флуоресцентная микроскопия. а б в ранняя стадия Межклеточное взаимодействие г поздняя стадия Для энтоза в суспензии необходимы контакты с высоким содержанием Е-кадгерина и βкатенина (Overholtzer et al., 2007). Мы исследовали взаимодействие клеток при энтозе в субстрат-зависимых культурах. Окрашивание клеток антителами к β-катенину выявило толстое кольцо этого белка между внедрившейся клеткой и энтозной вакуолью на первой стадии энтоза, что говорит о большом количестве контактов между наружной и внутренней клетками. Начиная с третьей стадии, данные контакты не обнаруживались. Кроме того, в работе методом ТЭМ впервые обнаружены специализированные контакты – десмосомы – между мембранами внедрившейся клетки и энтозной вакуоли. При окрашивании клеток антителами к плакоглобину на первой стадии энтоза выявлены плотно прилегающие друг к другу десмосомы, расположенные по периферии внутренней клетки. Затем их количество уменьшалось, и впоследствии они не обнаруживались. Нами была выдвинута гипотеза, что сильная адгезия между клетками необходима для удержания внедрившейся клетки внутри формирующейся энтозной вакуоли. Выход же внутренней клетки из наружной, который 9 происходит в некоторых случаях (Overholtzer and Brugge, 2008), вероятно, может быть связан с отсутствием таких контактов. Актиновый цитоскелет и его роль при энтозе Внедрение клетки в клетку в суспензии происходит за счёт работы акто-миозинового комплекса во внедряющейся клетке (Overholtzer et al., 2007). В субстрат-зависимых культурах по нашим данным в этом процессе также участвуют актиновые филаменты энтозной клетки, обеспечивающие образование псевдоподии (Insall and Machesky, 2009). В связи с этим мы исследовали роль микрофиламентов при энтозе. Для этого клетки обрабатывали цитохалазином В. Подсчёт энтотического индекса показал, что воздействие ингибитором привело к снижению числа энтозных клеток в культуре (Рисунок 4а). частота встречаемости энтозов (%) а Воздействие цитохалазином В приводит к уменьшению числа внедрившихся клеток, отмывка от данного ингибитора увеличивает число энтозов клеток линии А431. а – гистограмма частоты встречаемости энтозов при воздействии цитохалазином В и после его отмывки; б – отмывка от цитохалазина В в течение 7 ч, образование кратера; в – отмывка в тече24 ч 48 ч 24 ч 48 ч 1 ч контроль б Рисунок 4. Актиновый цитоскелет цитохалазин В в 4ч 7 ч 16 ч 19 ч 24 ч 48 ч ние 16 ч, накрывание внедряющейся отмывка от цитохалазина В клетки псевдоподией наружной клет- г ки; г – отмывка в течение 24 ч, структура “клетка-в-клетке”; б-г – СЭМ. Красные стрелки указывают на внедрившиеся клетки, зелёные – 10 µm 10 µm 10 µm на наружные. Начиная с 8 ч инкубации клеток с ингибитором, не выявлялись ранние стадии энтоза, через 48 ч визуализировались почти полностью деградировавшие внедрившиеся клетки. Эффект от воздействия цитохалазином В был обратим. После отмывки от ингибитора клетки вновь стали вступать в энтоз (Рисунок 4 б-г). В итоге энтотический индекс достиг контрольного значения через 24 ч после отмывки от цитохалазина В (Рисунок 4а). Таким образом, при энтозе требуется полимеризация актина в обеих клетках-участницах (во внедряющейся для продавливания плазматической мембраны энтозной клетки за счёт акто-миозинового сокращения (Sun et al., 2014), а в наружной для образования псевдоподии). Данный эксперимент позволяет синхронизировать начало вступления клеток в процесс энтоза. 10 Система микротрубочек и её вклад в энтоз При энтозе в суспензии микротрубочки регулируют упругость клеток (Xia et al., 2014). Для того, чтобы установить вклад микротрубочек в процесс энтоза на субстрате, мы обрабатывали культуру клеток А431 нокодазолом. Через 30 мин все микротрубочки в клетках были деполимеризованы. В ходе 24-часового воздействия число энтозных клеток постепенно уменьшалось вплоть до нулевого значения (Рисунок 5а). Через 8 и более часов инкубации с нокодазолом не обнаруживались ранние стадии энтоза. Методом СЭМ были выявлены лишь клетки, способные образовывать кратер в энтозной клетке. Однако наружная клетка при этом не формировала псевдоподию (Рисунок 5б). а б частота встречаемости энтозов (%) Воздействие нокодазолом на клетки приводит к уменьшению числа энтозов в культуре 0 1 3 5 8 15 19 24 Время инкубации с нокодазолом (ч) ! Рисунок 5. Микротрубочки в клетках линии А431. а – гистограмма частоты встречаемости энтозов в культуре при воздействии нокодазолом; б – обработка клеток нокодазолом в течение 15 ч, СЭМ. Таким образом, в процессе внедрения одной клетки в другую участвуют как актиновый цитоскелет, так и микротрубочки. Роль последних при энтозе в субстрат-зависимых культурах, по-видимому, заключается не только в регулировании упругости клеток, но и в обеспечении доставки дополнительных мембранных везикул к плазматической мембране наружной клетки для увеличения её поверхности, что необходимо при формировании кратера и псевдоподии. Локализация аппарата Гольджи при энтозе На ранних стадиях энтоза аппарат Гольджи внедрившейся клетки располагается компактно вблизи ядра. На поздних стадиях визуализируются расширенные цистерны и везикулы аппарата Гольджи, хаотично располагающиеся в цитоплазме. В наружной клетке на ранних стадиях энтоза аппарат Гольджи перераспределяется и располагается по периферии энтозной вакуоли. На четвёртой/пятой стадии происходит повторное перераспределение диктиосом в наружной клетке – они возвращаются в околоядерную область (Рисунок 6 а, б). 11 а в б 10 µm г д ! ае ж б вз иг д к м ж нз !и о к п 10 µm ле ! Рисунок 6. Мембранные органеллы клеток. а, б – аппарат Гольджи клеток линии А431: а – первая стадия энтоза; б – пятая стадия; окрашивание клеток антителами к белку аппарата Гольджи 58К и цитохимическое окрашивание ядер с помощью DAPI, флуоресцентная микроскопия. в-д – Аутофагосомы клеток линии НаСаТ: в – первая стадия; г – вторая стадия; д – пятая стадия; прижизненное окрашивание клеток моноданзилкадаверином, флуоресцентная микроскопия. Красными стрелками показаны аутофагосомы внедрившейся клетки, зелёными – наружной. л-п – Лизосомы клеток линии А431: е, л – первая стадия; ж, м – вторая стадия; з, н – третья стадия; и, о – четвёртая стадия; к, п – пятая стадия; е-к – фазовоконтрастная микроскопия; л-п – прижизненное окрашивание акридиновым оранжевым, флуоресцентная микроскопия. Аутофагосомы В ходе работы обнаружено, что при энтозе происходит активация аутофагии. На первой стадии данного типа КК во внедрившейся клетке выявляются единичные аутофагосомы. На второй и третьей стадиях их количество увеличивается, на четвёртой стадии они сливаются, а на пятой стадии моноданзилкадаверином окрашивается вся клетка. В наружной клетке на первой стадии обнаружены немногочисленные хаотично расположенные аутофагосомы. На второй стадии они локализуются по периферии энтозной вакуоли. На четвёртой стадии выявляются крупные аутофагосомы вокруг энтозной вакуоли (Рисунок 6 в-д). Кислый везикулярный компартмент При энтозе в суспензии внедрившиеся клетки, как правило, подвергаются лизосомоопосредованной деградации (Overholtzer et al., 2007). Мы также выявили активацию кислого 12 компартмента в клетках при энтозе на субстрате. Во внедрившейся клетке на первой стадии энтоза немногочисленные лизосомы локализуются вблизи ядра. На второй стадии их количество увеличивается, они обнаруживаются по периферии клетки, а на третьей стадии распределяются по всей цитоплазме. На четвёртой стадии лизосомы сливаются, образуя крупные везикулы, происходит закисление цитоплазмы клетки, а на пятой стадии акридиновым оранжевым окрашивается вся энтозная вакуоль (Рисунок 6 л-п). В наружной клетке лизосомы распределяются по периферии энтозной вакуоли на первой и второй стадиях. В дальнейшем они локализуются вблизи ядра, либо рассредоточены по цитоплазме клетки. Их количество различно вне зависимости от стадии энтоза. На основе наших результатов и данных литературы мы составили схему, иллюстрирующую последовательность событий, связанных с модификацией мембраны энтозной вакуоли и лизосомо-опосредованной деградацией внутренней клетки (Рисунок 7). а г в б д е Рисунок 7. Схема последовательности событий, происходящих после внедрения клетки в клетку. а – Локализация клеточных органелл и состав мембраны энтозной вакуоли сразу после внедрения (аналогичен составу плазматической мембраны); б – активация белков аутофагии и первая модификация мембраны энтозной вакуоли данными белками; в – полная замена плазматической мембраны энтозной вакуоли на мембрану аутофагосом; г – активация лизосом и вторая модификация мембраны энтозной вакуоли мембранными белками лизосом; д – сформировавшаяся мембрана энтозной вакуоли; е – закисление энтозной вакуоли и деградация внедрившейся клетки. Обозначения: АГ – аппарат Гольджи; БА – белки аутофагии; ВК – внутренняя клетка; Л – лизосома; М – митохондрия; НК – наружная клетка; ЭВ – энтозная вакуоль; Я – ядро. 13 После внедрения клетки в клетку первая оказывается внутри вакуоли, мембрана которой представляет собой плазматическую мембрану наружной клетки. Затем происходит первая модификация мембраны энтозной вакуоли за счёт белков аутофагии (Florey et al., 2011). С такой модифицированной мембраной могут сливаться лизосомы наружной клетки, дополнительно образующиеся вокруг энтозной вакуоли за счёт работы аппарата Гольджи, также располагающегося в этой области (Rohrer and Kornfeld, 2001). В результате мембранный белок лизосом LAMP1 оказывается в составе мембраны энтозной вакуоли, защищая наружную клетку от лизосомных гидролаз (Fukuda, 1991), высвобождающихся в полость энтозной вакуоли. Параллельно происходит активация продукции лизосом во внутренней клетке. Следовательно, деградация внедрившейся клетки происходит за счёт активации процессов аутофагии и кислого везикулярного компартмента в обеих клетках-участницах энтоза. Энтоз – энергозатратный процесс. Энергия в клетках генерируется в результате работы митохондрий. В связи с этим мы исследовали состояние этих мембранных органелл при энтозе и обнаружили, что митохондрии внедрившейся клетки сохраняют потенциал на мембране вплоть до третьей стадии энтоза. Митохондрии наружной клетки меняют локализацию, в основном располагаясь вокруг энтозной вакуоли. Роль аппарата Гольджи при энтозе Мы предполагаем, что основные функции аппарата Гольджи при энтозе – это: 1) участие в везикулярном транспорте, который, вероятно, обеспечивает увеличение площади поверхности плазматической мембраны энтозной клетки (Holthuis and Levine, 2005) (необходимое для образования кратера и псевдоподии); 2) участие в образовании лизосом. Для того, чтобы проверить, действительно ли аппарат Гольджи играет важную роль при энтозе, мы обрабатывали клетки линии А431 брефелдином А после синхронизации их вступления в энтоз с помощью предварительной 48-часовой обработки цитохалазином В. Были обнаружены единичные энтозные клетки на всех сроках эксперимента по сравнению с контрольными клетками (отмытыми после воздействия цитохалазином В) (Рисунок 8а). Анализ полученных препаратов с помощью СЭМ выявил структуры, когда одна клетка сильно распластана по поверхности другой клетки или ошаренная клетка продавливает плазматическую мембрану прикреплённой клетки, в результате чего формируется кратер (Рисунок 8 б, в). Однако при этом не выявлено накрывание ошаренной клетки плазматической мембраной прикреплённой клетки за счёт формирования псевдоподии последней. Нарушение работы аппарата Гольджи вследствие обработки культуры брефелдином А так же, как и его дезорганизация, вызванная нокодазолом (Cole et al., 1996), не блокировали деградацию ранее внедрившейся клетки за счёт уже имеющихся лизосом. Таким образом, 14 нами было впервые показано, что аппарат Гольджи необходим для внедрения клетки в клетку. Его роль в процессе деградации внедрившейся клетки не столь существенна. Brefeldin decreases the of ВоздействиеAбрефелдином А наnumber клетки приводит к уменьшению числа энтозов internalized cells в культуре Cell internalization rate частота встречаемости энтозов (%) (%) а Рисунок 8. Воздействие брефелдином А на клетки линии А431. 1,5 а – гистограмма частоты встречае- 1 мости энтозов при культивирова- 0,5 нии клеток с брефелдином А (пос- 0 К C 5,5h 5,5 БФА C К BFA 15h 5,5 15 5,5h БФА C 19h К BFA 15 19 15h БФА C 24h К BFA 19 24 19h БФА BFA 24 24h Время инкубации (ч) К – контроль; БФА – с брефелдином А клеток с помощью цитохалазина В); б, в – обработка клеток брефелдином А в течение 8 и 19 ч, СЭМ. в б ле предварительной синхронизации Оранжевые стрелки указывают на клетки, расположенные сверху на прикреплённых к субстрату клетках, зелёные – на клетки, прикреплённые к субстрату. Судьба внедрившейся клетки при энтозе Внедрившаяся клетка при энтозе, как правило, подвергается деградации. Однако не установлено, как завершается данный тип КК. В связи с этим мы провели прижизненное наблюдение за клетками линии MCF7 (Рисунок 9). 0ч 9ч Рисунок 19 ч 9. жизненное дение за Принаблю- судьбой внедрившейся клетки линии MCF7. DIC. Красная стрелка 10 µm 28 ч указывает на внед- 48 ч 78 ч рившуюся клетку. ! 15 В начале наблюдения обнаружена внедрившаяся клетка без видимых морфологических изменений, находящаяся внутри крупной вакуоли (первая стадия энтоза). Наружная клетка хорошо распластана и имеет выраженную ламеллу. Через 9 ч внутренняя клетка поджимается, а энтозная становится менее распластанной (вторая стадия). Через 19 ч наблюдается деградация внедрившейся клетки: размер значительно уменьшается, ядро не выявляется (пятая стадия). Через 28 ч уменьшается размер энтозной вакуоли. Через 48 ч происходит экструзия непереваренного материала, ядро наружной клетки вновь становится овальным. Через 78 ч энтозная клетка хорошо распластана по субстрату. Обнаружен также альтернативный вариант судьбы внедрившейся клетки, а именно – её выход из энтозной клетки. В литературе описаны редкие случаи, когда внедрившаяся клетка при КК деградирует по механизму апоптоза (Wang et al., 2009). В связи с этим мы окрашивали клетки антителами к активной каспазе-3, цитохрому с и проводили прижизненное окрашивание на активные формы кислорода – основные маркёры апоптоза. Не было выявлено окрашивание ни одним из перечисленных способов. Это означает, что механизм апоптоза не включён в процесс деградации внедрившейся клетки. Выявление гетеротипического энтоза В многоклеточных организмах опухолевые и нормальные клетки находятся в непосредственной близости. Для того, чтобы установить, может ли между ними происходить взаимодействие, приводящее к гетеротипическому энтозу, мы провели эксперименты по а б сокультивированию нормальных кератиноцитов человека (НаСаТ) с опухолевыми клетками (А431 и MCF7). Также были поставлены эксперименты по сокультивированию клеток двух опухолевых линий для сравнения их энтотической активности. При постановке экспериментов одну из опухолевых клеточных линий метили бромдезоксиуридином. Через 2-е суток инкубирования клеток с BrdU процент меченых клеток в культуре А431 составлял 99-100%, в культуре МСF7 – 95-99%. Экперименты проводили по схемам, представленным на рисунке 10. а а а в б бб в в в Рисунок 10. Схемы постановки экспериментов по сокультивированию клеток двух культур в разных комбинациях. Меченые BrdU клетки – зелёные, немеченые – фиолетовые. 16 10 µm в в бгб д е Для того, чтобы проверить, влияет ли исходное состояние клеток, суспензионное или аа прикреплённое, на энтотический индекс, мы сокультивировали меченую и немеченую популяции клеток линии А431 по описанным схемам. Было выявлено, что изначальное µmв клетку при 1010µm 10 µm состояние клеток не оказывает существенного влияния на внедрение клетки аа гомотипическом энтозе. зее и При сокультивировании клеток разных линий мы обнаружили 4 типа взаимодействия между ними: немеченая клетка в немеченой и меченая в меченой – гомотипический энтоз, а также немеченая в меченой и меченая в немеченой – гетеротипический энтоз (Рисунок 11). 1010µm µm 10 µm 10 µm а в ж вдвд гббгб а 10 µm Рисунок 11. Энтоз при гззее к л ии м сокультивировании клеток разных линий. Немеченые клетки – НаСаТ, 10 µm 10 µm г дбг а еддв ж ж к ж кзезе 1010µm µm ! лзл ии 1010µm µm 10 µm меченые клетки – MCF7. мм а-г – фазово-контрастная микроскопия; д-з – окрашивание клеток антитела- д жж 10µm µm 10 10 µm 10 µm г ! ! ккззе ли и л 10 µm 10 µm мм ми к 10 BrdU, µm флуорес- 10 µm ! центная микроскопия. ! Сокультивирование клеток линий HaCaT и А431. В эксперименте I (Рисунок 10а) было обнаружено достоверное различие между двумя вариантами гетеротипического энтоза: 10 µm 10 µm 10 µmлинии НаСаТ, 10 µm клетки линии А431 чаще внедрялись в клетки чем наоборот (Рисунок 12а). В 10 µm ! ! л м ! ли ккз эксперименте II (Рисунок 10б) наблюдалась аналогичная тенденция.! Это может быть связано с м ж к вв дд ж бб г тем, что клетки линии А431 обладают более высокой энтотической активностью по сравнению с клетками линии НаСаТ. Сокультивирование10клеток линий10НаСаТ и MCF7. В эксперименте II достоверно µm µm 10 µm ! ! ! показано, что клетки в клетки линии MCF7, чем наоборот л ! м линии НаСаТ чаще внедряются (Рисунок 12б). В эксперименте III (Рисунок 10в) такая тенденция сохраняется. Вероятно, клетки линии НаСаТ, изначально находясь в суспензионном состоянии, “легче” внедряются в другие клетки, т.к. им не требуется предварительно открепляться от субстрата. Помимо этого, 10 µm в культуре MCF7 можно выделить! две популяции клеток: 1) крупные и хорошо распластанные; 2) небольшого размера и значительно менее распластанные. Таким образом, клетки линии MCF7, относящиеся к первой популяции, занимали большую площадь поверхности, в результате чего увеличивалась вероятность взаимодействия с ними внедряющихся клеток. 17 ! Сами же эти клетки, по-видимому, редко внедрялись в другие клетки из-за своих крупных размеров. ! в б средний % клеток к общему числу энтозов средний % клеток к общему числу энтозов а медианы 25%-75% выпавшие значения * ◊ * • ◊ • НаСаТ А431 A431 НаСаТ НаСаТA431А431 НаСаТ внутри внутри внутри внутри внутри внутри внутри НаСаТ внутри A431 A431 НаСаТ А431 НаСаТ НаСаТ А431 !! медианы 25%-75% выпавшие значения • • НаСаТ MCF-7 НаСаТ MCF-7 НаСаТMCF7 MCF7 НаСаТ НаСаТ MCF7 MCF7 НаСаТ внутри внутри внутри внутри внутри внутри внутри внутри внутри внутри НаСаТ MCF-T MCF-7 НаСаТ НаСаТвнутри MCF7 внутри MCF7 НаСаТ НаСаТ MCF7 MCF7 НаСаТ ! MCF7 А431 MCF7 А431 внутри внутри внутри внутри MCF7 А431 А431 MCF7 Рисунок 12. Гистограммы распределения различных вариантов энтозов при сокультивировании клеток разных линий. а – НаСаТ и А431, статистически значимые различия между группами: * - гомотипический и гетеротипический энтоз (р = 0,05), • - НаСаТ внутри А431 и А431 внутри НаСаТ (р = 0,0209), ◊ - НаСаТ внутри НаСаТ и внутри А431 (р = 0,05); б – НаСаТ и MCF7, p = 0,0209; в – А431 и MCF7, статистически значимые различия между группами: * - гомотипический и гетеротипический энтоз (p = 0,0079); ∆ - MCF7 внутри MCF7 и MCF7 внутри А431 (р = 0,0209). Сокультивирование опухолевых клеток линий MCF7 и А431. Показано, что клетки линии MCF7 чаще внедряются в клетки той же линии, нежели в клетки линии А431 (Рисунок 12в). Достоверное отличие между клетками линий А431 и MCF7 по способности внедряться друг в друга отсутствует. Однако во всех экспериментах клетки культуры А431 демонстрировали тенденцию чаще внедряться в клетки линии MCF7, чем наоборот. Можно предположить, что опухолевые клетки различаются по способности внедряться в клетки других линий. Это может быть связано с тем, что клетки линии А431 имеют меньший размер и являются примером более “аггрессивного” рака, чем клетки линии MCF7. Таким образом, во всех вариантах сокультивирования клеток разных линий обнаружен гетеротипический энтоз. Это свидетельствует о том, что для внедрения клетки в клетку не требуется наличие специфических молекул узнавания, присущих клеткам одной линии. Тем не менее, из полученных результатов видно, что гомотипический энтоз преобладает над гетеротипическим. Мы объясняем это тем, что клетки одной культуры более адгезивны друг к другу, чем к клеткам другой линии (Balzer and Konstantopoulos, 2012). Кроме того, часть гомотипических энтозов могла формироваться ещё до начала эксперимента и сохраняться в течение 24-х ч, т.к. энтоз длится более 20 ч (Overholtzer and Brugge, 2008 и рисунок 9). 18 Деградация внедрившейся клетки при гетеротипическом энтозе осуществляется за счёт работы кислого везикулярного компартмента обеих клеток-участниц. Заключение В работе на субстрат-зависимых культурах клеток человека впервые описаны процессы, включённые в реализацию энтоза, а именно: формирование десмосом между клеткамиучастницами, увеличение площади поверхности наружной клетки, необходимое для формирования структуры “клетка-в-клетке”, активация аутофагии и лизосом в наружной и внутренней клетках, обеспечивающая деградацию последней. В дальнейшем предстоит охарактеризовать молекулярные механизмы осуществления данных процессов. Это позволит управлять энтозом. Тот факт, что КК присущ большому спектру опухолей, ставит вопрос о роли энтоза в развитии опухолей – способствует ли он опухолевой прогрессии (при увеличении степени малигнизации опухолей в них наблюдается усиление энтотической активности) или выступает как фактор супрессии опухолей. Решение данной проблемы в дальнейшем может позволить использование данного варианта клеточной гибели в противоопухолевой терапии. В ходе исследования для клеток линии А431 показано, что внедрение одной клетки в другую происходит за счёт работы актинового цитоскелета обеих клеток-участниц энтоза. Получены новые данные о роли микротрубочек и аппарата Гольджи наружной клетки в данном процессе, обеспечивающих увеличение площади её поверхности. В дальнейшем необходимо детализировать роль аппарата Гольджи в ходе энтоза. Нами впервые выявлено присутствие большого количества десмосом между внутренней и наружной клетками. Изучение роли десмосом при энтозе, вероятно, позволит открыть новые функции данных контактов, т.к. на сегодняшний день известно, что их основная задача – обеспечение интеграции клеток внутри ткани (Kottke et al., 2006). В работе впервые предложено разделение энтоза на 5 стадий после формирования структуры “клетка-в-клетке”. Это может использоваться для систематизации его описания. В дальнейшем предстоит выделить и подробно охарактеризовать больше стадий энтоза, начиная с процесса внедрения клетки в клетку. Нами представлен комплексный метод верификации локализации одной клетки внутри другой (корреляционная микроскопия). Впервые описан способ синхронизации вступления клеток в энтоз, который может быть применим впоследствии для изучения механизмов каждой стадии энтоза. Всё это имеет важное практическое методологические подходы к изучению энтоза. 19 значение, т.к. предоставляет В работе показано, что при сокультивировании клеток разных линий гомотипический энтоз, как правило, преобладает над гетеротипическим. Внедряющиеся клетки могут быть как опухолевыми, так и нормальными. В связи с этим целесообразно будет исследовать биологическую роль обоих вариантов энтоза. Сравнение результатов экспериментов по сокультивированию нормальных клеток с опухолевыми клетками двух линий выявило, что опухолевые клетки различаются по способности вступать в гетеротипический энтоз с нормальными клетками. В связи с этим предстоит более детально исследовать, существуют ли конкурентные взаимоотношения опухолевых клеток различного тканевого происхождения, оказавшихся в контакте с нормальными клетками, посредством энтоза. Так, биологическая роль данного типа КК может быть связана с популяционным контролем клеток одного или разных типов: с помощью энтоза может осуществляться отбор более приспособленных опухолевых или нормальных клеток. В ходе дальнейшей работы также предстоит ответить на вопросы о причинах, инициирующих внедрение клетки в клетку, о механизмах узнавания между клетками и о том, что определяет судьбу внедрившейся клетки. Подводя итог, можно заключить, что энтоз вызывает интерес как с точки зрения фундаментальной науки о клетках (в качестве примера клеток, выполняющих неканонические функции), так и с точки зрения прикладной биологии (КК может являться мишенью для борьбы с различными опухолями). Выводы 1. В субстрат-зависимых культурах клеток эпителиального происхождения – нормальных клеток НаСаТ и опухолевых клеток А431 и MCF7 – выявлен гомотипический вариант энтоза. 2. На основе морфологических изменений клеток-участниц энтоза данный процесс разделён на 5 стадий. 3. Разработан метод синхронизации вступления клеток в процесс энтоза в культуре. 4. На ранней стадии энтоза между внедрившейся и энтозной клетками линии А431 впервые обнаружено большое количество десмосом. 5. В ходе энтоза происходит разрушение адгезивных контактов между внутренней и наружной клетками, в том числе и десмосом, а также перераспределение мембранных органелл и реорганизация цитоскелета в энтозной клетке. 6. На клетках линии А431 показано, что для внедрения одной клетки в другую необходима работа актинового цитоскелета, микротрубочек и аппарата Гольджи. 20 7. При совместном культивировании нормальных клеток с опухолевыми, а также опухолевых клеток разного типа друг с другом наблюдается как гомотипический, так и гетеротипический энтоз. При этом: а) гомотипический энтоз, как правило, преобладает над гетеротипическим; б) опухолевые клетки различаются по способности вступать в гетеротипические взаимодействия с нормальными клетками: клетки линии А431 обладают бóльшей способностью внедряться в нормальные кератиноциты человека, чем клетки линии MCF7. 8. При гомо- и гетеротипическом энтозе внедрившаяся клетка, как правило, подвергается деградации, в процессе которой происходит активация лизосом в обеих клетках-участницах. Для гомотипического энтоза также показана активация процессов аутофагии. СПИСОК ОПУБЛИКОВАННЫХ РАБОТ Статьи в журналах 1. A.S. Garanina and M.A. Savitskaya. Cell Cannibalism in Human Epidermoid Carcinoma A431 Cell Line. // Moscow University Biological Sciences Bulletin – 2013 – Vol. 68, No. 1 – pp. 5–7. Original Russian Text A.S. Garanina, M.A. Savitskaya, 2013, published in Vestnik Moskovskogo Universiteta. Biologiya – 2012 – No. 4 – pp. 3–7; 2. A.S. Garanina, L.A. Khashba, and G.E. Onishchenko. Stages of Cell Cannibalism – Entosis – in Normal Human Keratinocyte Culture. // Biochemistry – 2015 – Vol. 80, No. 11 – pp. 1469-1477. Original Russian Text A.S. Garanina, L.A. Khashba, G.E. Onishchenko, 2015, published in Biokhimiya – 2015 – Vol. 80, No. 11 – pp. 1688-1697. Тезисы конференций 1. Гаранина А.С. Клеточный каннибализм в культуре клеток эпидермоидной карциномы человека А431. // Материалы XVIII международной научной конференции студентов, аспирантов и молодых ученых «Ломоносов – 2011», серия Биология – 2011 – с. 280-281; 2. Гаранина А.С., Савицкая М.А., Онищенко Г.Е. Роль цитоскелета в процессе клеточного каннибализма в культуре клеток эпидермоидной карциномы человека А431. // Цитология – 2011 – том 53, серия 9 – с. 698; 3. Garanina A.S., Milovanova N.V., Onishchenko G.E. Heterotypic cell cannibalism in normal and neoplastic cell lines of epithelial origin. // Cell Technology Week 2013: abstracts book of the I International Scientific Conference of Students and PhD Students – 2013 – p. 38, Kyiv, Ukraine; 21 4. Гаранина А.С., Онищенко Г.Е. Нарушение целостности элементов цитоскелета ингибирует процессы клеточного каннибализма в культивируемых клетках А431. // Онтогенез – 2013 – том 44, №4 – с. 225; 5. Гаранина А.С., Онищенко Г.Е. Цитоскелет и везикулярный транспорт необходимы для внедрения одной клетки в другую, но не для ее деградации в процессе энтоза. // Материалы международной научной конференции «Актуальные вопросы морфогенеза в норме и патологии» – 2014 – с. 73-75; 6. Милованова Н.В., Гаранина А.С., Онищенко Г.Е. Гетеротипический клеточный каннибализм в культурах нормальных и опухолевых клеток эпителиального происхождения. // Материалы международной научной конференции «Актуальные вопросы морфогенеза в норме и патологии» – 2014 – с. 170-172; 7. Милованова Н.В., Гаранина А.С. Взаимодействие нормальных и опухолевых клеток эпителиального происхождения при сокультивировании: гетеротипический клеточный каннибализм in vitro. // Материалы XXI международной научной конференции студентов, аспирантов и молодых ученых «Ломоносов – 2014», серия Биология – 2014 – с. 161-162. 22