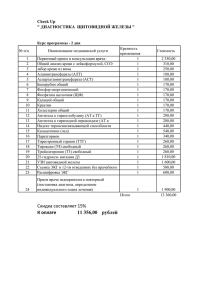
017265 B1 017265 B1 (11) 017265
реклама
 017265")
Евразийское патентное ведомство (19) (13) B1 ОПИСАНИЕ ИЗОБРЕТЕНИЯ К ЕВРАЗИЙСКОМУ ПАТЕНТУ (45) Дата публикации и выдачи патента 2012.11.30 Номер заявки 200870231 (22) Дата подачи заявки 2007.02.03 (54) (51) Int. Cl. C12Q 1/68 (2006.01) C12P 19/34 (2006.01) C12N 15/63 (2006.01) A61K 38/00 (2006.01) C07K 1/00 (2006.01) C07H 21/02 (2006.01) C07H 21/04 (2006.01) A01N 43/04 (2006.01) ПРИМЕНЕНИЕ АНТИТЕЛА IMC-A12, КОТОРОЕ ЯВЛЯЕТСЯ ИНГИБИТОРОМ IGF-IR, ДЛЯ ЛЕЧЕНИЯ РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ (56) US-20050164970 YU, H. et al., Role of the insulin-like growth factor family in cancer development and progression, J. Natl. Cancer Insti., Vol. 92, No. 18, September 2000, pages 1472-1489, esp 1472-3, section entitled "IGF Receptors", pages 1473-1475; section entitled IGFs and Cancer Experimental Evidence, pages 1475-6 WO-9825642 Изобретатель: (74) Представитель: (57) Изобретение относится к способу лечения рака предстательной железы с помощью терапии депривацией андрогенов и антагониста рецептора инсулиноподобного фактора роста (IGF-IR). Хотя уровень ответа рака предстательной железы на терапию депривацией андрогенов (ADT) является высоким, выжившие раковые клетки неизбежно становятся андрогеннезависимыми (AI) и опухолевый рост продолжается. В изобретении ингибируют или задерживают переход от андрогензависимого рака к андрогеннезависимому раку, значительно снижая риск возникновения рецидива и улучшая результат лечения. Людвиг Дейл, Плаймейт Стефен Р. (US) Медведев В.Н. (RU) B1 (72) 017265 (31) 60/765,072 (32) 2006.02.03 (33) US (43) 2009.12.30 (86) PCT/US2007/003164 (87) WO 2007/092453 2007.08.16 (71)(73) Заявитель и патентовладелец: ИМКЛОУН ЭЛЭЛСИ; ЮНИВЕРСИТИ ОФ ВАШИНГТОН (US) B1 017265 (12) (21) 017265 (11) 017265 Государственное финансирование Настоящее изобретение частично осуществлено при поддержке правительства США в рамках гранта № СА85859 от National Institutes of Health и гранта № W81XWH-04-1-0912 от Department of Defense. Соответственно, правительство США обладает определенными правами на это изобретение. Перекрестная ссылка на родственные заявки По заявке на данное изобретение испрашивается приоритет заявки США № 60/765072, поданной 3 февраля 2006 г., которая представлена в описании в качестве ссылки в полном объеме. Область изобретения Настоящее изобретение относится к способу лечения рака предстательной железы посредством терапии депривацией андрогенов и антагониста рецептора инсулиноподобного фактора роста (IGF-IR). Способ ингибирует или задерживает переход андрогензависимого рака в андрогеннезависимый рак, значительно снижает риск возникновения рецидива и улучшает исход лечения. Предпосылки изобретения Рак предстательной железы является наиболее распространенным раком некожного происхождения и является второй наиболее часто встречаемой причиной смертности от рака у мужчин в США. Большинство случаев рака предстательной железы вначале является андрогензависимыми (AD). Клеткам рака предстательной железы вначале необходимы андрогены для продолжения пролиферации. Ответ на абляцию тестостерона посредством терапии депривацией андрогенов (ADT) или хирургически (орхиэктомия), или медикаментозно (агонисты GnRH или эстрогены) приводит к быстрой индукции апоптоза в чувствительных клетках рака предстательной железы. Положительный уровень ответа составляет приблизительно 86%, исходя из снижения уровня специфического антигена предстательной железы (PSA) и стабилизации или снижения объема опухоли. Гибель клеток, как правило, происходит в пределах от первых нескольких суток до недели. Однако за положительным ответом следует период остановки роста, при котором остающиеся клетки обладают тенденцией не погибать. Спустя 18-36 месяцев после гормональной абляции рост возобновляется в 90% случаев. Во всех случаях выжившие раковые клетки становятся андрогеннезависимыми или нечувствительными, и следует андрогеннезависимый (AI) рост опухоли. Так как ADT вначале является очень эффективной, то может принести большую пользу терапия, которая может обладать преимуществами ADT и продлевать или усиливать ее эффекты. Как представляется, независимость от андрогенов возникает посредством множества механизмов. Мутации в гене рецептора андрогенов редко диагностируют, но их частота увеличивается после воздействия антиандрогена флутамида. Однако эти мутации не возникают у большинства пациентов и не объясняют большинство случаев гормонрезистентного заболевания. Высокие уровни bcl-2 наблюдают с большей частотой при запущенном заболевании по сравнению с локализованным заболеванием. Таким образом, способность индуцировать апоптоз снижается при прогрессировании заболевания. Пролиферация клеток, несущих мутации в гене опухолевого супрессора р53, утрата рецепторов TGF- и экспрессия пептидных факторов роста, вероятно, играет роль в развитии гормонрезистентного состояния. Однако эти процессы не объясняют скорость и частоту развития. Рецептор инсулиноподобного фактора роста (IGF-IR) представляет собой повсеместно распространенный трансмембранный рецептор тирозинкиназы, который необходим для нормального роста и развития плода и постнатального роста и развития. IGF-IR может стимулировать пролиферацию клеток, дифференцировку клеток, изменения размера клеток и защищать клетки от апоптоза. Его также рассматривали в качестве квазинеобходимого для трансформации клеток (рассмотрено в Adams et al., Cell. Mol. Life Sci. 57:1050-93 (2000); Baserga, Oncogene 19:5574-81 (2000)). IGF-IR расположен на поверхности клеток большинства типов клеток и служит в качестве сигнальной молекулы для факторов роста IGF-I и IGF-II (вместе обозначаемых в дальнейшем IGF). IGF-IR также связывается с инсулином, хотя и с величиной аффинности, на три порядка меньшей по сравнению с его связыванием с IGF. IGF-IR представляет собой преформированный гетеротетрамер, содержащий две альфа- и две бета-цепи, ковалентно связанные посредством дисульфидных связей. Субъединицы рецептора синтезируются в виде части одиночной полипептидной цепи 180 кДа, которая затем протеолитически процессируется на альфа- (130 кДа) и бета(95 кДа) субъединицы. Вся альфа-цепь является внеклеточной и содержит участок для связывания лиганда. Бета-цепь содержит трансмембранный домен, тирозинкиназный домен и С-концевой участок, который необходим для дифференцировки и трансформации клеток, но является несущественным для опосредованной митогеном передачи сигнала и защиты от апоптоза. IGF-IR обладает очень сильным сходством с рецептором инсулина (IR), в частности, в пределах последовательности бета-цепи (70% гомология). Благодаря этой гомологии в недавних исследованиях показали, что эти рецепторы могут образовывать гибриды, содержащие один димер IR и один димер IGF-IR (Pandini et al., Clin. Cane. Res. 5:1935-19 (1999)). Образование гибридов происходит как в нормальных, так и трансформированных клетках, и содержание гибрида зависит от концентрации двух гомодимерных рецепторов (IR и IGF-IR) в клетке. В одном исследовании 39 образцов рака молочной железы, хотя как IR, так и IGF-IR сверхэкспрессировались во всех образцах опухоли, содержание гибридного рецептора постоянно превышало уровни обоих гоморецепторов приблизительно в 3 раза (Pandini et al., Clin. Cane. -1- 017265 Res. 5:1935-44 (1999)). Хотя гибридные рецепторы состоят из пар IR и IGF-IR, гибрид связывается селективно с IGF, с аффинностью, сходной с аффинностью IGF-IR, и только слабо связывается с инсулином (Siddle and Soos, The IGF System. Humana Press, p. 199-225. 1999). Эти гибриды, таким образом, могут связываться с IGF и передавать сигналы как в нормальных, так и в трансформированных клетках. Эндокринная экспрессия IGF-I регулируется преимущественно гормоном роста и происходит в печени, но последние данные позволяют предположить, что многие другие типы тканей также способны экспрессировать IGF-I. Этот лиганд, таким образом, подвергается эндокринной и паракринной регуляции, а также аутокринной в случае многих типов опухолевых клеток (Yu, H. and Rohan, J., J. Natl. Cancer Inst. 92:1472-89 (2000)). Рецептор андрогенов (AR) состоит из 3 функциональных и структурных доменов: N-концевого (модуляторного) домена; ДНК-связывающего домена (депозитарный № Interpro IPR001628), который опосредует специфическое связывание с последовательностями ДНК-мишени (лиганд-чувствительные элементы); и гормонсвязывающего домена. N-концевой домен (NTD) является уникальным для рецептора андрогенов и охватывает приблизительно первые 530 остатков; высококонсервативный ДНКсвязывающий домен меньше (приблизительно 65 остатков) и занимает центральную часть белка; и лигандсвязывающий домен (LBD) для гормона расположен на С-конце рецептора. В отсутствие лиганда рецепторы стероидного гормона, как предполагают, слабо связаны с компонентами ядра; связывание гормона значительно усиливает аффинность рецептора. Взаимоотношение между рецептором андрогенов (AR), андрогеном и раком предстательной железы является комплексным. На распределение AR между ядром и цитоплазмой влияет андроген и удаление андрогена. Например, иммунореактивность AR наблюдают только в ядрах клеток LuCaP 35, растущих у интактных самцах мышей, однако сильную иммунореактивность наблюдают в цитоплазме и ядрах LuCaP 35, растущих у интактных самцах мышей и впоследствии кастрированных. Сущность изобретения Изобретение относится к лечению андрогензависимых опухолей, таких как рак предстательной железы. Опухоли предстательной железы, как правило, стимулируются андрогенами, такими как тестостерон, и обладают андрогензависимым (AD) ростом. Таким образом, лечение рака предстательной железы, как правило, включает терапию, при которой происходит прекращение действия андрогена на клетки рака предстательной железы. Однако в значительной доле случаев рака предстательной железы в конечном итоге происходит переход к независимости от андрогенов (AI). Обнаружено, что введение антагониста IGF-IR в сочетании с терапией посредством андрогенной депривации (ADT) ингибирует или предотвращает переход AD-опухолей в AI-опухоли. Соответственно, изобретение относится к способу лечения андрогензависимого рака путем проведения андрогенной депривации и введения антагониста IGF-IR. В варианте осуществления по изобретению андрогензависимый рак является раком предстательной железы. Согласно изобретению антагонист IGF-IR может представлять собой внеклеточный антагонист или внутриклеточный антагонист, и можно применять более одного антагониста. В более широком смысле, изобретение относится к ингибированию обусловленной IFG-IR передачи сигнала и модулированию компонента пути таким образом, чтобы ингибировать переход опухолевых клеток от AD к AI. Внеклеточные антагонисты включают в качестве неограничивающих примеров белки или другие биологические молекулы, которые связываются с IGF-IR или его лигандом (IGF). В некоторых вариантах осуществления изобретения внеклеточный антагонист ингибирует связывание IGF-IR с IGF. В одном из вариантов осуществления связывающийся белок представляет собой антитело, такое как, например, IMC-A12. В другом варианте осуществления связывающийся белок является растворимым лигандсвязывающим фрагментом IGF-IR. Внутриклеточные антагонисты IGF-IR могут представлять собой биологические молекулы, но обычно являются малыми молекулами. В варианте осуществления изобретения антагонист IGF-IR является малой молекулой, выбранной из AG1024, NVP-AEW541 и BMS-554417. Эффективность различных антагонистов в ингибировании обусловленной IGF-IR передачи сигнала можно наблюдать, например, путем анализа состояния компонентов пути обусловленной IGF-IR передачи сигнала. В одном из вариантов осуществления ингибирование IGF-IR наблюдают по снижению фосфорилирования Akt. В другом варианте осуществления ингибирование обусловленной IGF-IR передачи сигнала наблюдают по снижению экспрессии сурвивина или -пептида тубулина (TUBB) . Антагонист IGF-IR по изобретению применяют с любой формой ADT. В варианте осуществления изобретения ADT включает орхиэктомию. В другом варианте осуществления изобретения ADT включает введение аналога рилизинг-фактора лютеинизирующего гормона. В другом варианте осуществления ADT включает введение антиандрогена. В другом варианте осуществления вводят ингибитор андрогена надпочечников. Согласно изобретению можно сочетать два или более способа ADT. Изобретение далее относится к ингибированию передачи сигнала посредством Akt. Соответственно, изобретение относится к введению модуляторов передающих сигнал белков, которые активируют Akt. В одном из вариантов осуществления такой модулятор представляет собой антагонист EGFR. -2- 017265 Согласно изобретению антагонист IGF-IR вводят в качестве адъюванта для ADT. В одном из вариантов осуществления ADT и введение антагониста IGF-IR начинают приблизительно в одно и то же время. В другом варианте осуществления сначала начинают ADT, и антагонист IGF-IR вводят перед тем, как андрогеннезависимый рак станет андрогеннезависимым. Изобретение далее относится к применению противоопухолевых средств совместно с ADT и введением антагониста IGF-IR. В варианте осуществления изобретения антагонист IGF-IR и средство ADT применяют вместе в качестве неоадъюванта для хирургического или радиационного лечения рака предстательной железы. Изобретение также относится к композициям, содержащим антагонист IGF-IR и средство ADT в лекарственной форме. Краткое описание чертежей На фиг. 1 представлено исследование, в котором изучали подкожные ксенотрансплантаты LuCap35 у мышей SCID. Всех мышей кастрировали, когда средний размер опухоли достигал 400 мм3. В контрольной группе мышам производили только кастрацию. В двух других группах вводили три раза в неделю IMC-A12, начиная через одну или две недели после кастрации. На фиг. 2 представлены уровни PSA у кастрированных контрольных мышей и у кастрированных мышей, которых лечили IMC-А12, начиная через одну (раннее начало) или две (позднее начало) недели после кастрации. На фиг. 3 представлено распределение рецептора андрогенов (AR) в ответ на стимуляцию IGF-IR посредством IGF и/или ингибирование IGF-IR посредством IMC-A12. Уровни AR в цитоплазме и ядре анализировали вестерн-блоттингом. На фиг. 4 представлен эффект антагониста IGF-IR (IMC-А12) на распределение рецептора андрогенов (AR) в андрогензависимых ксенотрансплантатах опухолей из клеток LuCaP35 у интактных мышей (левая колонка) и андрогеннезависимых ксенотрансплантатах опухолей из клеток LuCaP35V у кастрированных мышей (правая колонка). На фиг. 5 представлена корреляция между количественным показателем AR и объемом опухоли. r=0,66, р<0,01. Величины для только кастрированных животных представлены светлыми кругами и величины для кастрированных + А12 при раннем и позднем начале лечения представлены темными кругами. Величины представляют собой среднюю величину для 100 ядер, распределенных в опухоли. На фиг. 6 представлены изменения экспрессии генов между двумя периодами времени для подкожных обработанных посредством А12 опухолей. Из 3170 уникальных генов на чипе с достаточным количеством данных для тестирования 21 подвергался активации (включая многие регулируемые андрогенами, обозначенные с помощью "*") и 41 подвергался подавлению с 10% добротности в позднем периоде времени, когда опухоли начинали рецидивировать, по сравнению с ранним периодом времени. На фиг. 7А представлена корреляция между показателем количества копий сурвивина и объемом опухоли (r=0,66, р0,01). На фиг. 7В представлена корреляция между показателем количества копий бета-пептида 3 тубулина и объемом опухоли (r=0,59, р0,01). Величины для только кастрированных представлены светлыми кругами и величины для кастрированных + А12 при раннем и позднем начале представлены темными кругами, каждая величина является средней величиной для трех осуществлений PCR. Подробное описание изобретения Обнаружено, что ингибиторы IGF-IR пригодны в терапии для лечения рака предстательной железы. В частности, введение антагониста IGF-IR в сочетании с терапией депривацией андрогенов (ADT) приводит к улучшению результата лечения по сравнению только с ADT. Обнаружено, что андрогены активируют экспрессию рецептора инсулиноподобного фактора ростаI и могут делать рак предстательной железы чувствительным к эффектам IGF-I. Сходным образом, переход к независимости от андрогенов, который наблюдают в клетках рака предстательной железы, может происходить в результате приспособлений клетки, при которых происходит усиление обусловленной рецептором андрогенов передачи сигнала, таких как увеличение уровней AR, которые делают клетку чувствительной к низким уровням циркулирующего андрогена, или мутации в AR, которые дают возможность активации с помощью неандрогенных стероидов. В самом деле, данные говорят о том, что обусловленная IGF-I передача сигнала может действительно опосредовать транслокацию AR в ядро в опухолевых клетках и приводить к активации AR-зависимых генов. Таким образом, предположили, что IGF-I может способствовать превращению андрогензависимого рака предстательной железы в андрогеннезависимый после терапии гормональной абляцией, способствуя обусловленной AR передаче сигнала в отсутствие циркулирующего андрогена. Последние данные для мужчин и для ксенотрансплантатов предстательной железы человека также говорят о том, что существующие в настоящее время способы андрогенной абляции не способны снижать количество андрогенов предстательной железы до уровней, которые более не вызывают активации рецептора андрогенов. Предстательная железа может действительно обладать способностью синтезировать DHT из нескольких стероидов-предшественников и, возможно, ацетата. -3- 017265 Таким образом, отсюда следует, что ингибирование обусловленной IGF-I передачи сигнала в дополнение к терапии гормональной абляцией может предотвращать превращение или увеличивать время до превращения рака предстательной железы в андрогеннезависимое заболевание, значительно задерживая возникновение рецидива. Антагонисты IGF-IR, таким образом, могут представлять собой эффективную адъювантную терапию по отношению к способам андрогенной депривации для лечения недавно диагностированного и местно развивающегося или метастатического гормонзависимого рака предстательной железы. Применение антагонистов IGF-IR в сочетании с андрогенной абляцией также обладает способностью блокировать опосредованное IGF возвращение из апоптоза. Механизмы, посредством которых IGFIR может предотвращать апоптоз, включают ингибирование киназы ras-raf-map, киназы PI3, включая mTOR, и опосредованную forkhead передачу сигнала, и 14-3-3. Другой механизм, посредством которого ингибирование IGF-IR может продлевать эффекты андрогенной абляции, осуществляется посредством поддержания опухоли в состоянии остановки клеточного цикла вслед за началом апоптоза. В предыдущих исследованиях показано, что антагонисты IGF-IR могут обладать положительным эффектом при использовании для лечения ксенотрансплантатов как андрогензависимых, так и андрогеннезависимых видов рака предстательной железы. Рост ксенотрансплантатов, хотя и замедленный, нельзя остановить или обратить. В настоящее время обнаружено, что антагонисты IGF-IR, в частности, пригодны для лечения рака предстательной железы при введении совместно с терапией депривацией андрогенов (ADT). Как правило, опухоли предстательной железы переходят к независимости от андрогенов и становятся нечувствительными к ADT. Как обнаружено ранее, такие андрогеннечувствительные опухоли обладают тенденцией не давать сильные ответы на антагонисты IGF-IR. Однако, как представлено в данной работе, время для прогрессии опухолей предстательной железы от AD до AI значительно увеличивается при терапии, которая сочетает ADT с введением антагониста IGF-IR. В течение этого более длительного периода опухоли уменьшаются в размере, и снижаются уровни PSA. Сочетанная терапия снижает высокий риск возникновения рецидива, который наблюдают при применении только ADT, и снижает риск развития метастазов рака. Лечение антагонистами IGF-IR также обладает преимуществами при лечении запущенного рака предстательной железы, при котором потенциально присутствуют или диагностированы метастазы. В моделях, в которых вводят клетки рака предстательной железы, наблюдают, что транслокация AR из цитоплазмы в ядро индуцируется не только стимуляцией андрогеном, но также, хотя и в меньшей степени, стимуляцией IGF-IR. Даже в присутствии андрогена, транслокация AR в присутствии андрогена и IGF снижается посредством антагониста IGF-IR. В предстательной железе после кастрации все еще детектируют низкие уровни андрогенов. Также сообщалось, что экспрессия IGF-IR, который участвует в передаче сигнала через Akt, сначала снижается в ответ на кастрацию, но затем усиливается, и далее стимуляция Akt этим фактором роста усиливает обусловленную AR передачу сигнала в ответ на низкие уровни андрогена. Как показано в данной работе, лечение антагонистом IGF-IR значительно задерживает восстановление роста опухолей у кастрированных мышей. Более того, существует хорошая корреляция между снижением AR в ядре и уменьшением объема опухоли. Это позволяет предположить, что ингибирование обусловленной IGF-IR передачи сигнала играет значительную роль в ингибировании обусловленной AR прогрессии опухоли. В описанных в данной работе экспериментах обусловленную IGF-IR передачу сигнала ингибируют с использованием антитела, обозначенного как А12, которое связывается с IGF-IR. В предыдущих экспериментах с использованием А12 и сходных антител показано, что происходит снижение фосфорилирования (т.е. активации) различных участвующих в передаче сигнала молекул, включая ERK и MAPK, в частности Akt. Эффект ингибирования IGF-IR обнаружен во множестве типов опухолевых клеток, включая линию опухоли предстательной железы М12 (Wu, J.D. et al. , 2005, Clin. Cancer Res. 11:3065-74) и клетки рака молочной железы MCF7 (Burtrum, D. et al., 2003, Cancer Res. 63:8912-21). Таким образом, следует принимать во внимание, что ту же самую или сходную адъювантную активность, наблюдаемую в данной работе для антагониста IGF-IR, можно наблюдать для средств, которые обладают таким же или сходным эффектом на активацию Akt. Показано, что лечение антагонистом IGF-IR приводит к ингибированию транслокации AR в ядро. Ингибирование можно наблюдать гистохимически или флуоресцентной микроскопией, а также по снижению уровней экспрессии индуцируемых AR генов. Два гена, связанные с устойчивостью к кастрации, сурвивин и -пептид тубулина, регулируются IGF-IR посредством активации Akt. Экспрессия генов супрессирована у кастрированных мышей, которых подвергали лечению антагонистом IGF-IR, по сравнению с теми, которых только кастрировали. Сходные ингибиторные эффекты на транслокацию AR и экспрессию генов, активированных Akt, можно наблюдать в ответ на специфичный в отношении Akt ингибитор или антагонист другого пути передачи сигнала, в который в значительной степени вовлечена Akt. Множество антагонистов IGF-IR можно применять согласно изобретению. Антагонисты IGF-IR могут представлять собой внеклеточные антагонисты или внутриклеточные антагонисты. Внеклеточные и внутриклеточные антагонисты IGF-IR могут являться биологическими молекулами, малыми молекулами или любым другим веществом, которое ингибирует активацию IGF-IR, например, посредством взаимо-4- 017265 действия с внеклеточной связывающей областью рецептора (т.е. внеклеточный антагонист), посредством ингибирования фосфорилирования внутриклеточного тирозинкиназного домена IGF-IR, или ингибированием взаимодействия с активацией любого другого клеточного компонента, вовлеченного в обусловленный IGF-IR путь передачи сигнала, таким образом, в конечном итоге ингибируя активацию генов или клеточную пролиферацию. В варианте осуществления настоящего изобретения внеклеточный антагонист IGF-IR взаимодействует с областью рецептора, связывающей внеклеточный лиганд, посредством достаточного физического или химического взаимодействия между антагонистом и внеклеточной связывающей областью рецептора таким образом, что блокируется связывание IGF-IR со своим лигандом (IGF), и ингибируется тирозинкиназная активность рецептора. Специалист в данной области примет во внимание, что примеры таких химических взаимодействий, которые включают ассоциацию или образование связи, известны в данной области и включают образование ковалентной связи, ионной связи, водородной связи и т.п. между антагонистом и внеклеточной связывающей областью. В варианте осуществления изобретения внеклеточный антагонист IGF-IR является биологической молекулой. Биологические молекулы включают в качестве неограничивающих примеров антитела или фрагменты антител, которые связываются с IGF-IR. В другом варианте осуществления антагонист IGF-IR может являться малой молекулой, которая блокирует связывание лиганда с IGF-IR. В другом варианте осуществления внеклеточный антагонист является веществом, которое блокирует лиганды IGF-IR или вызывает их деградацию. Один из примеров представляет собой растворимый внеклеточный фрагмент IGF-IR, который связывается с IGF. Другой пример такого вещества представляет собой IGF-связывающий белок (IGFBP), такой как, например, IGFBP-1, IGFBP-2 и IGFBP-3, который может связываться с IGF, таким образом ограничивая активацию рецептора IGF. В другом варианте осуществления изобретения низкомолекулярный ингибитор связывается с лигандсвязывающим доменом IGF-IR и блокирует с его помощью связывание лиганда IGF-IR и активацию рецептора. Несмотря на нежелание вдаваться в теоретические подробности, принято считать, что внеклеточный антагонист IGF-IR ингибирует все каскады передачи сигнала, инициируемые конформационными изменениям во внеклеточной области IGF-IR после активации IGF-IR. Это ингибирование включает IGFIR на поверхности, а также те IGF-IR, которые интернализованы внутри клетки. Например, предполагают, что тирозинкиназы активированного рецептора (RTK) могут подвергаться интернализации в эндосому посредством покрытой кларитином ямки, при этом все еще сохраняя активность в передаче сигнала. После интернализации такие рецепторы или возвращаются на поверхность клетки, или подвергаются деградации в эндосоме или лизосоме. Другой путь ингибирования опосредованной IGF-IR передачи сигнала представляет собой подавление экспрессии IGF-IR. В варианте осуществления изобретения антагонист IGF-IR связывается с рецептором и способствует интернализации и деградации рецептора. В другом варианте осуществления антагонист IGF-IR снижает экспрессию рецептора. Биологические молекулы, в контексте настоящего изобретения, включают все аминокислоты, нуклеотиды, липиды и полимеры моносахаридов, которые, как правило, обладают молекулярной массой более 650 Да. Таким образом, биологические молекулы включают, например, олигопептиды, полипептиды, пептиды и белки, олигонуклеотиды и полинуклеотиды, такие как, например, ДНК и РНК, и олигосахариды и полисахариды. Биологические молекулы дополнительно включают производные любых из вышеописанных молекул. Например, производные биологических молекул включают липиды и гликозилированные производные или олигопептиды, полипептиды, пептиды и белки. Производные биологических молекул дополнительно включают липидные производные олигосахаридов и полисахаридов, например липополисахариды. Чаще всего, биологические молекулы являются антителами или их функциональными производными. Малые молекулы включают органические соединения, такие как гетероциклы, пептиды, сахариды, стероиды и т.п., металлоорганические соединения, соли органических соединений и металлоорганических соединений и неорганические соединения. Атомы в малых молекулах связаны вместе посредством ковалентной и ионной связей; первая является типичной для малых органических соединений, таких как низкомолекулярные ингибиторы тирозинкиназы, и последняя является типичной для малых неорганических соединений. Расположение атомов в малой органической молекуле может представлять собой цепь, например углерод-углеродную цепь или цепь из углерода и гетероатомов, или может представлять собой кольцо, содержащее атомы углерода, например бензольную или полициклическую систему, или сочетание из углерода и гетероатомов, т.е. гетероциклы, такие как пиримидин или хиназолин. Хотя малые молекулы могут обладать любой молекулярной массой, они, как правило, включают молекулы, которые могут в других отношениях рассматриваться как биологические молекулы, за исключением того, что их молекулярная масса не превышает 650 Да. Малые молекулы включают как встречающиеся в природе соединения, такие как гормоны, нейротрансмиттеры, нуклеотиды, аминокислоты, сахара, липиды и их производные, так и соединения, полученные синтетически, или традиционным органическим синтезом, биологически опосредованным синтезом, или их сочетанием. См., например, Ganesan, Drug Discov. Today 7(1):47-55 (Jan. 2002); Lou, Drug Discov. Today, 6(24):1288-1294 (Dec. 2001). Соединения можно -5- 017265 модифицировать для усиления эффективности, стабильности, фармацевтической совместимости и т.п. Внутриклеточные антагонисты IGF-IR могут представлять собой биологические молекулы, такие как мутантные субъединицы рецептора, внутриклеточные связывающие белки (например, внутриклеточно экспрессируемые фрагменты антител) и т.п. В предпочтительном варианте осуществления внутриклеточные антагонисты являются малыми молекулами. Низкомолекулярные ингибиторы включают в качестве неограничивающих примеров малые молекулы, которые модифицируют или блокируют ATPсвязывающий домен, субстратсвязывающие области или киназный домен IGF-IR. Низкомолекулярные ингибиторы также включают вещества, которые являются ингибиторами других компонентов пути обусловленной IGF-IR передачи сигнала, включая в качестве неограничивающих примеров путь rasмитоген-активируемой протеинкиназы (MAPK) и путь киназы фосфатидилинозита-3 (PI3K)-Akt. Для идентификации антагонистов можно осуществлять скрининг низкомолекулярных библиотек на ингибиторную активность с использованием высокопроизводительных биохимических, ферментативных или клеточных анализов. Анализы можно разрабатывать для детектирования способности тестируемого соединения ингибировать связывание IGF-IR с лигандами IGF-IR или субстратом IRS-1 или ингибировать образование функциональных рецепторов из димеров IGF-IR. Внутриклеточный антагонист IGF-IR может ингибировать тирозинкиназную активность IGF-IR путем связывания с внутриклеточной областью, несущей киназный домен, или ингибированием ее активации, или путем связывания с любым внутриклеточным белком, вовлеченным в путь передачи сигнала от IGF-IR, или ингибированием его активации. Низкомолекулярные антагонисты IGF-IR включают, например, ингибиторы селективной киназы рецептора инсулиноподобного фактора роста-I NVP-AEW541 (Garcia-Echeverria, С. et al., 2004, Cancer Cell. 5:231-9) и NVP-ADW742 (Mitsiades, С. et al., 2004, Cancer Cell. 5:221-30), INSM-18 (Insmed Incorporated), которые избирательно ингибируют IGF-IR и HER2 и тирозинкиназные ингибиторы трифостины AG1024 и AG1034 (Parrizas, M. et al., 1997, Endocrinology. 138:1427-33), которые ингибируют фосфорилирование путем блокирования связывания субстрата и обладают значительно более низким IC50 для ингибирования фосфорилирования IFG-IR, по сравнению с фосфорилированием IR. Пикроподофиллин производное циклолигнана (РРР) представляет собой другой антагонист IGF-IR, который ингибирует фосфорилирование IGF-IR без нарушения активности IR (Girnita, A. et al., 2004, Cancer Res. 64:236-42). Другие низкомолекулярные антагонисты IGF-IR включают производные бензимидазола BMS-536924 (Wittman, M. et al., 2005, J. Med. Chem. 48:5639-43) и BMS-554417 (Haluska P. et al., 2006, Cancer Res. 66:362-71), которые ингибируют IGF-IR и IR почти в равной степени. Для соединений, которые ингибируют рецепторы в дополнение к IGF-IR, следует указать, что величины IC50, измеренные in vitro в прямых анализах связывания, могут не отражать величины IC50, измеренные ex vivo или in vivo (т.е. в интактных клетках или организмах). Например, там, где желательно избегать ингибирования IR, соединение, которое ингибирует IR in vitro, может не оказывать значительного влияния на активность рецептора при использовании in vivo в концентрации, при которой эффективно ингибируется IGF-IR. Антисмысловые олигодезоксинуклеотиды, антисмысловые РНК и малые ингибиторные РНК (миРНК) обеспечивают направленную деградацию мРНК, таким образом предотвращая трансляцию белков. Соответственно, можно ингибировать экспрессию тирозинкиназ рецепторов и других белков, играющих ключевую роль в обусловленной IGF передаче сигнала. Способность антисмысловых олигонуклеотидов супрессировать экспрессию генов обнаружили более 25 лет назад (Zamecnik and Stephenson, 1978, Proc. Natl. Acad. Sci. USA. 75:280-84). Антисмысловые олигонуклеотиды спариваются с мРНК и пре-мРНК и могут потенциально нарушать несколько стадий процессинга РНК и трансляции матричной РНК, включая сплайсинг, полиаденилирование, экспорт, стабильность и трансляцию белка (Sazani and Kole, 2003, J. Clin. Invest. 112:481-86). Однако две наиболее мощные и широко используемые антисмысловые стратегии представляют собой деградацию мРНК или пре-мРНК посредством РНКазы Н и изменение сплайсинга путем нацеливания на аберрантные точки сплайсинга. РНКаза Н распознает гетеродуплексы ДНК/РНК и расщепляет РНК приблизительно посередине между 5'- и 3'-концами олигонуклеотида ДНК. Ингибирование IGF-IR с помощью антисмысловых олигонуклеотидов представлено в Wraight, Nat. Biotechnol. 18:521-6. Природные РНК-опосредованные механизмы могут регулировать стабильность мРНК, трансляцию матричной РНК и организацию хроматина (Mello and Conte, 2004, Nature. 431:338-42). Более того, экзогенно введенная длинная двухцепочечная РНК (дцРНК) является эффективным инструментом для подавления транскрипции генов у множества низших организмов. Однако у млекопитающих длинные дцРНК вызывают реакции, обусловленные высокой токсичностью, которые связаны с эффектами вирусной инфекции и продукцией интерферона (Williams, 1997, Biochem. Soc. Trans. 25:509-13). Чтобы этого избежать, Elbashir и сотрудники (Elbashir, et al., 2001, Nature. 411:494-98) положили начало использованию миРНК, образованных 19-мерными дуплексами с 5'-фосфатами и 3'-выступающими концами из 2 оснований на каждой цепи, которые при введении в клетки избирательно деградируют мРНК-мишени. Действие интерферирующей дцРНК у млекопитающих, как правило, включает две ферментативные стадии. Сначала Dicer, фермент РНКаза III-типа, расщепляет дцРНК на 21-23-мерные сегменты миРНК. Затем РНК-индуцируемый ингибирующий комплекс (RISC) расплетает дуплекс РНК, спаривается с комплементарной областью смысловой мРНК на одной цепи и инициирует расщепление в участке из 10 -6- 017265 нуклеотидов выше 5'-конца цепи миРНК (Hannon, 2002, Nature. 418:244-51). В кратком изложении, химически синтезированные миРНК в диапазоне 19-22-мерных молекул не нуждаются в стадии с участием Dicer и могут входить в комплекс RISC непосредственно. Следует указать, что любая цепь дуплекса РНК может потенциально входить в комплекс RISC, но композиция олигонуклеотидов может влиять на выбор цепей. Таким образом, для достижения избирательной деградации конкретной мРНК-мишени дуплекс должен способствовать входу в комплекс компонента антисмысловой цепи путем наличия относительно слабого спаривания на своем 5'-конце (Khvorova, 2003, Cell 115:209-16). Экзогенные миРНК можно получить в виде синтезированных олигонуклеотидов или экспрессировать с плазмидных или вирусных векторов (Paddison and Hannon, 2003, Curr. Opin. Mol. Ther. 5:217-24). В последнем случае молекулыпредшественники обычно экспрессируются в виде коротких шпилечных РНК (shPHK), содержащих петли из 4-8 нуклеотидов и стебли из 19-30 нуклеотидов; они затем расщепляются с помощью Dicer с образованием функциональных миРНК. Антитела анти-IGF-IR для применения согласно настоящему изобретению обладают одним или несколькими из следующих свойств. 1) Антитела связываются с внешним доменом IGF-IR и ингибируют связывание IGF-I или IGF-II с IGF-IR. Ингибирование можно определять, например, анализом прямого связывания с использованием очищенного или связанного с мембраной рецептора. В данном варианте осуществления антитела по настоящему изобретению или их фрагменты предпочтительно связываются с IGF-IR, по меньшей мере, также сильно, как и природные лиганды IGF-IR (IGF-I и IGF-II). 2) Антитела нейтрализуют IGF-IR. Связывание лиганда, например IGF-I или IGF-II, с внешним, внеклеточным доменом IGF-IR стимулирует аутофосфорилирование бета-субъединицы и фосфорилирование субстратов IFG-IR, включал MAPK, Akt и IRS-I. Нейтрализация IGF-IR включает ингибирование, снижение, инактивацию и/или нарушение одной или нескольких из этих активностей, в норме связанных с передачей сигнала. Нейтрализацию можно определить in vivo, ex vivo или in vitro с использованием, например, тканей, культивируемых клеток или очищенных клеточных компонентов. Нейтрализация включает ингибирование гетеродимеров IGF-IR/IR, а также гомодимеров IGF-IR. Таким образом, нейтрализация IGF-IR обладает различными эффектами, включая ингибирование, снижение, инактивацию и/или нарушение роста (пролиферации и дифференцировки), ангиогенез (образования кровеносных сосудов, инвазии и метастазирования) и клеточную подвижность и метастазирование (адгезии клеток и инвазивности). Одним их способов измерения нейтрализации IGF-IR является ингибирование тирозинкиназной активности рецептора. Ингибирование тирозинкиназы можно определить с использованием хорошо известных способов, например измерением уровня аутофосфорилирования рекомбинантного киназного рецептора и/или фосфорилирования природных или синтетических субстратов. Таким образом, анализы фосфорилирования пригодны для определения нейтрализующих антител в контексте настоящего изобретения. Фосфорилирование можно детектировать, например, в анализе ELISA или вестерн-блоттинге с использованием антитела, специфичного к фосфотирозину. Некоторые анализы тирозинкиназной активности описаны в Panek et al., 1997, J. Pharmacol. Exp. Thera. 283:1433-44 и Batley et al., 1998, Life Sci. 62:143-50. Антитела по изобретению вызывают снижение фосфорилирования тирозина в IGF-IR по меньшей мере приблизительно на 75%, предпочтительно по меньшей мере приблизительно на 85% и более предпочтительно по меньшей мере приблизительно на 90% в клетках, которые реагируют на лиганд. Другим способом измерения нейтрализации IGF-IR является ингибирование фосфорилирования расположенных ниже субстратов IGF-IR. Соответственно, можно измерять уровень фосфорилирования MAPK, Akt или IRS-I. Снижение фосфорилирования субстрата составляет по меньшей мере приблизительно 50%, предпочтительно по меньшей мере приблизительно 65%, более предпочтительно по меньшей мере приблизительно 80%. Кроме того, для определения нейтрализации IGF-IR можно использовать способы детекции экспрессии белка, в которых измеряемые белки регулируются тирозинкиназной активностью IGF-IR. Примером такого белка, который связан с прогрессированием рака и устойчивостью к лекарственным средствам, является сурвивин, который является членом семейства ингибиторов апоптоза (IAP). Хотя регуляция сурвивина сложна и опосредована более чем одним путем, была показана регуляция, опосредованная Akt и усиливаемая IGF-1. См., например, Zhang et al., 2005, Oncogene, 24:2474-82. Способы анализа экспрессии генов включают иммуногистохимию (IHC) для детекции экспрессии белка, флуоресцентную гибридизацию in situ (FISH) для детекции амплификации генов, анализы конкурентного связывания радиолиганда, способы блоттинга на основе твердых матриц, такие как нозерн- и саузерн-блоттинги, полимеразная цепная реакция после обратной транскрипции (RT-PCR) и ELISA. См., например, Grandis et al., 1996, Cancer, 78:1284-92; Shimizu et al., 1994, Japan J. Cancer Res., 85:567-71; Sauter et al., 1996, Am. J. Path., 148:1047-53; Collins, 1995, Glia 15:289-96; Radinsky et al., 1995, Clin. Cancer Res. 1:19-31; Petrides et al., 1990, Cancer Res. 50:3934-39; Hoffmann et al., 1997, Anticancer Res. 17:4419-26; Wikstrand et al., 1995, Cancer Res. 55:3140-48. -7- 017265 Анализы ex vivo также можно использовать для определения нейтрализации IGF-IR. Например, ингибирование рецептора тирозинкиназы можно исследовать в митогенных анализах с использованием линий клеток, стимулированных лигандом рецептора в присутствии и в отсутствие ингибитора. Линия рака молочной железы MCF7 (American Type Culture Collection (ATCC), Rockville, MD) является такой линией клеток, которая экспрессирует IGF-IR и стимулируется IGF-I или IGF-II. Другой способ включает тестирование ингибирования роста опухолевых клеток, экспрессирующих IGF-IR, или клеток, трансфицированных, чтобы экспрессировать IGF-IR. Ингибирование можно также исследовать с использованием моделей опухолей, например инъецировать мыши опухолевые клетки человека. Антитела по настоящему изобретению не ограничены каким-либо конкретным механизмом нейтрализации IGF-IR. Антитела анти-IGF-IR по настоящему изобретению могут связываться с внешней стороны с клеточным поверхностным рецептором IGF-IR, блокировать связывание лиганда (например, IGF-I или IGF-II) и последующую передачу сигнала, опосредованную связанной с рецептором тирозинкиназой, и предотвращать фосфорилирование IGF-IR и других расположенных ниже белков в каскаде передачи сигнала. 3) Антитела подавляют IGF-IR. Количество IGF-IR, присутствующего на поверхности клетки, зависит от продукции, интернализации и деградации белка рецептора. Количество IGF-IR, присутствующего на поверхности клетки, можно измерять опосредованно, детектированием интернализации рецептора или молекулы, связанной с рецептором. Например, интернализацию рецептора можно измерять контактированием клеток, которые экспрессируют IGF-IR, с мечеными антителами. Связанные с мембраной антитела затем смывают, собирают и определяют их количество. Интернализованные антитела определяют лизированием клеток и детектированием метки в лизатах. Другой способ состоит в прямом измерении количества рецептора, присутствующего на клетке, после обработки антителами анти-IGF-IR или другим веществом, например, при помощи анализа флуоресцентно-активированным клеточным сортингом клеток, окрашенных на поверхностную экспрессию IGFIR. Окрашенные клетки инкубируют при 37°С и интенсивность флуоресценции измеряют с течением времени. В качестве контроля часть окрашенной популяции можно инкубировать при 4°С (условия, при которых интернализация рецептора останавливается). IGF-IR на клеточной поверхности можно детектировать и измерять с использованием другого антитела, которое специфично к IGF-IR и которое не блокирует тестируемое антитело или конкурирует с ними за связывание (Burtrum, et al., 2003, Cancer Res. 63:8912-21). Обработка клетки, экспрессирующей IGF-IR, антителом по изобретению приводит к снижению количества IGF-IR на клеточной поверхности. В предпочтительном варианте осуществления снижение в ответ на обработку антителом по изобретению составляет по меньшей мере приблизительно 70%, более предпочтительно по меньшей мере приблизительно 80% и еще более предпочтительно по меньшей мере приблизительно 90%. Значительное снижение можно наблюдать уже через 4 ч. Другое измерение подавления представляет собой снижение общего количества рецепторного белка, присутствующего в клетке, и отражает деградацию внутренних рецепторов. Соответственно, обработка клеток (в частности, раковых клеток) антителами по изобретению приводит к снижению общего клеточного IGF-IR. В предпочтительном варианте осуществления снижение составляет по меньшей мере приблизительно 70%, более предпочтительно по меньшей мере приблизительно 80% и еще более предпочтительно по меньшей мере приблизительно 90%. Для лечения человека антитела представляют собой предпочтительно антитела человека, но могут также являться гуманизированными или химерными антителами. Одним из предпочтительных антител человека, которое связывается с IGF-IR, является А12 (см. WO 2005016970). Другим предпочтительным антителом человека является 2F8 (см. WO 2005016970). Пригодные антитела дополнительно включают антитела анти-IGF-IR, которые конкурируют с IMC-A12 или IMC-2F8 за связывание с IGF-IR, a также антитела, которые связываются с другими эпитопами (т.е. антитела, которые связываются с другими эпитопами и обладают свойствами, как описано ранее, такими как блокирование лиганда, интернализация рецептора и т.д., но не конкурируют с IMC-A12 или IMC-2F8). Другие неограничивающие примеры нейтрализующих антител анти-IGF-IR, пригодных согласно изобретению, описаны в Wang et al. (WO 2003/1000008; US 2004/0018191) и Singh et al. (WO 2003/106621; US 2003/0235582). Нуклеотидные и аминокислотные последовательности нескольких упомянутых в данной работе антител представлены в табл. 1. -8- 017265 Таблица 1 SEQ ID NO вариабельных доменов антител и CDR (нуклеотид/аминокислота) Антитела, которые можно использовать согласно изобретению, включают целые иммуноглобулины, антигенсвязывающие фрагменты иммуноглобулинов, а также антигенсвязывающие белки, которые содержат антигенсвязывающие домены иммуноглобулинов. Антигенсвязывающие фрагменты иммуноглобулинов включают, например, Fab, Fab' и F(ab')2. Получены другие типы антител, которые сохраняют специфичность связывания, но обладают другими характеристиками, которые могут оказаться желательными, включая биспецифичность, поливалентность (более двух участков связывания), компактный размер (например, только связывающие домены). Одноцепочечные антитела содержат два вариабельных домена при отсутствии некоторых или всех константных доменов целых антител, из которых они получены. Таким образом, с их помощью можно решать некоторые проблемы, связанные с использованием целых антител. Например, одноцепочечные антитела обладают тенденцией не принимать участия в некоторых нежелательных взаимодействиях между константными областями тяжелых цепей и другими биологическими молекулами. Кроме того, одноцепочечные антитела обладают значительно меньшим размером по сравнению с целыми антителами и могут обладать лучшей проникающей способностью, чем целые антитела, позволяя одноцепочечным антителам более эффективно локализоваться на участках связывания антигена-мишени и связываться с ними. Более того, относительно малый размер одноцепочечных антител снижает вероятность вызывания ими нежелательного иммунного ответа у реципиента по сравнению с целыми антителами. Полимерные одноцепочечные антитела, где каждая одиночная цепь обладает одним VH- и одним VL-доменом, ковалентно связанным с первым пептидным линкером, можно ковалентно связать по меньшей мере одним или несколькими пептидными линкерами с образованием поливалентных одноцепочечных антител, которые могут являться моноспецифическими или полиспецифическими. Каждая цепь поливалентного одноцепочечного антитела включает вариабельный фрагмент легкой цепи и вариабельный фрагмент тяжелой цепи и связана пептидным линкером по меньшей мере с одной другой цепью. Пептидный линкер состоит по меньшей мере из 15 аминокислотных остатков. Максимальное количество аминокислотных остатков составляет приблизительно одну сотню. Два одноцепочечных антитела можно объединить с образованием димерного антитела, также известного как бивалентный димер. Димерные антитела обладают двумя цепями и двумя участками связывания и могут являться моноспецифическими или биспецифическими. Каждая цепь димерного антитела содержит VH-домен, соединенный с VL-доменом. Домены связаны с линкерами, которые являются достаточно короткими для предотвращения спаривания между доменами одной цепи, таким образом осуществляя спаривание между комплементарными доменами разных цепей для восстановления двух антигенсвязывающих участков. Сходным образом, три одноцепочечных антитела можно объединить с образованием тримерного антитела, также известного как тривалентный тример. Тримерные антитела конструируют таким образом, что аминокислотный конец VL- или VH-домена непосредственно слит с карбоксильным концом VL- или VH-домена (т.е. без какой-либо линкерной последовательности). Тримерные антитела могут являться моноспецифическими, биспецифическими или триспецифическими. Также получены биспецифические антитела, которые являются бивалентными в отношении каждого антигенсвязывающего участка. Например, в Zhu (WO 01/90192) описано антитело с четырьмя связывающими участками, которое в других отношениях обладает структурой и сохраняет эффекторные функции существующего в природе антитела. В Zhu (WO 2006/020258) описано биспецифическое антитело, которое содержит два димерных антитела и константные области Ig. Таким образом, антитела по изобретению и их фрагменты включают в качестве неограничивающих примеров природные антитела, бивалентные фрагменты, такие как (Fab')2, моновалентные фрагменты, такие как Fab, одноцепочечные антитела, одноцепочечные Fv (scFv), однодоменные антитела, поливалентные одноцепочечные антитела, димерные антитела, тримерные антитела и т.п., которые специфически связываются с антигенами. Антагонисты IGF-IR проиллюстрированы в данной работе на примере IMC-A12, моноклонального антитела человека, которое связывается с внеклеточным доменом IGF и блокирует связывание IGF. Свойства IMC-A12 и сходное антитело человека представлены в международной публикации WO 2005/016970. -9- 017265 Эффекты антагонистов IGF-IR по изобретению на клетки андрогензависимого рака предстательной железы включают один или несколько из следующего. 1) IGF может опосредовать активацию или транслокацию AR в отсутствие андрогена. Антагонисты IGF-IR по изобретению блокируют опосредованную IGF транслокацию. 2) Антагонисты IGF-IR опосредуют усиление уничтожения клеток или ингибирование пролиферации опухолевых клеток. 3) Снижается опосредованная AR экспрессия генов, активируемых рецептором андрогенов. Гены, обладающие опосредованной AR экспрессией, включают, например, PSA и TMPRSS2 (трансмембранная сериновая протеаза). Согласно изобретению антагонист IGF-IR вводят субъекту, обладающему раком предстательной железы в сочетании с терапией депривацией андрогенов (ADT; также называется гормональной терапией). Цель ADT состоит в снижении уровней мужских гормонов (андрогенов, таких как тестостерон) в организме. Андрогены, продуцируемые в основном в яичках, могут действительно стимулировать рост клеток рака предстательной железы. Снижение уровней андрогена обычно приводит к тому, что рак предстательной железы уменьшается в размере или растет более медленно. ADT применяют в нескольких случаях: в качестве терапии первой линии (начальной) пациентов, не подлежащих хирургическому вмешательству или радиационной терапии, или пациентов, которых невозможно излечить этими видами лечения вследствие того, что рак уже распространился за пределы предстательной железы; после начального лечения, такого как хирургия или радиационная терапия, если рак сохраняется или возвращается в исходное состояние; в качестве дополнения (адъюванта) к радиационной терапии как начальному лечению в определенных группах мужчин с высоким риском рецидивирования рака; и перед хирургическим вмешательством или радиационной терапией (неоадъювантная терапия) в попытке уменьшить размеры рака и сделать другой вид лечения более эффективным. Согласно изобретению антагонист IGF-IR вводят в сочетании с ADT в любой ситуации, где в других случаях можно применять ADT. Антагонист IGF-IR является адъювантом, который усиливает и/или пролонгирует эффект ADT. Существует несколько способов, применяемых в качестве ADT. Орхиэктомия включает удаление яичек, в которых продуцируется более 90% андрогенов, преимущественно тестостерона. При удалении этого источника в большинстве случаев рак предстательной железы уменьшается в размере. Хотя она является необратимой и приводит к множеству нежелательных побочных эффектов, как правило, связанных с изменением уровней гормонов в организме, орхиэктомия, вероятно, представляет собой наименее дорогостоящий и простейший способ снижения продукции андрогенов, и ее можно осуществить в виде простой амбулаторной процедуры. Аналоги рилизинг-фактора лютеинизирующего гормона (LHRH) (также называемые агонистами LHRH) снижают уровни тестостерона также эффективно, как и орхиэктомия, посредством снижения уровня андрогенов, преимущественно тестостерона, вырабатываемых яичками. Аналоги LHRH инъецируют или помещают под кожу в виде небольших имплантатов, и их вводят или ежемесячно или каждые 3, 4, 6 или 12 месяцев. Примеры аналогов LHRH включают леупролид, гозерелин и трипторелин. Возможные побочные эффекты аналогов LHRH сходны с побочными эффектами орхиэктомии, и они в значительной степени возникают вследствие изменений уровней гормонов. Антиандрогены блокируют способность организма использовать любые андрогены. Даже после орхиэктомии или в ходе лечения аналогами LHRH небольшое количество андрогенов все еще продуцируется надпочечниками. Лекарственные средства этого типа включают флутамид, бикалутамид и нилутамид. Эти лекарственные средства, как правило, принимают ежедневно в виде пилюль. Лечение антиандрогенами часто сочетают с орхиэктомией или лечением аналогами LHRH. Это сочетание называют комбинированной андрогенной блокадой (CAB). Более того, антиандроген можно добавлять, если лечение орхиэктомией или аналогом LHRH само по себе больше не дает эффекта. В нескольких последних исследованиях сравнивали эффективность антиандрогенов самих по себе с эффективностью агонистов LHRH. Большинство исследований не обнаружили отличий в уровнях выживаемости, но в нескольких исследованиях обнаружено, что антиандрогены незначительно менее эффективны. Побочные эффекты антиандрогенов у пациентов, которых уже лечили орхиэктомией или агонистами LHRH, как правило, не являются серьезными. Диарея представляет собой основной побочный эффект, хотя также могут встречаться тошнота, проблемы с печенью и усталость. Основное отличие от агонистов LHRH состоит в том, что антиандрогены обладают меньшим количеством связанных с сексуальными функциями побочных эффектов и позволяют поддерживать либидо и потенцию, если применять только их. Ингибиторы андрогенов надпочечников можно вводить вследствие того, что низкий уровень андрогенов, продуцируемых надпочечниками, может оказаться достаточным для обеспечения непрерывной стимуляции. После андрогенной абляции подмножество клеток рака предстательной железы может становиться гиперчувствительными к андрогенам, и надпочечники являются источником от 5 до 10% периферического тестостерона. Два наиболее общепринятых средства ингибирования продукции андрогенов надпочечниками являются аминоглютетимид и кетоконазол. - 10 - 017265 Другие примеры супрессирующих андрогены лекарственных средств включают диэтилстильбестрол (DES), мегестрола ацетат, ципротерона ацетат и преднизон. Эстрогены ранее являлись основной альтернативой орхиэктомии для мужчин с запущенным раком предстательной железы, но вследствие их возможных побочных эффектов, которые включают образование тромбов и увеличение молочных желез, эстрогены в значительной степени заменены аналогами LHRH и антиандрогенами. Согласно изобретению курс лечения антагонистом IGF-IR проводят начиная до, во время или после начала ADT. Курс введения антагонистом IGF-IR следует сочетать с ADT, но сочетание не обязательно является полным. Например, антагонист IGF-IR можно вводить в любое время в течение ремиссии, полученной в результате андрогенной абляции. В варианте осуществления изобретения антагонист IGF-IR вводят в пределах 24 месяцев после андрогенной абляции для лечения первичных или метастатических опухолей. В другом варианте осуществления антагонист IGF-IR вводят в пределах 18 месяцев после андрогенной абляции. В варианте осуществления изобретения антагонист IGF-IR вводят в течение или близко к концу периода гибели клеток, который наблюдают при лечении ADT, и он будет предотвращать или задерживать последующую пролиферацию AI-клеток. В варианте осуществления изобретения введение антагониста IGF-IR начинают в пределах двух недель после андрогенной абляции. В другом варианте осуществления введение начинают в пределах одной недели после андрогенной абляции. Антагонисты IGF-IR по изобретению можно вводить с антагонистами, которые нейтрализуют другие рецепторы, вовлеченные в опухолевый рост. Особенный интерес представляют рецепторы, вовлеченные в путь передачи сигнала, включающий Akt. Например, как предполагают, в передачу сигнала посредством EGFR или HER2 (erbB2) вовлечена активация Akt. Соответственно, антагонисты IGF-IR по изобретению можно сочетать с внутриклеточными или внеклеточными антагонистами EGFR или HER2. Антагонисты EGFR или HER2 включают антигенсвязывающие белки, которые связываются с внеклеточным доменом EGFR или HER2 и блокируют связывание одного или нескольких лигандов и/или нейтрализуют индуцированную лигандом активацию. Антагонисты также включают антитела или другие связывающие белки, которые связываются с лигандом EGFR и ингибируют связывание EGFR с лигандом. Лиганды EGFR включают, например, EGF, TGF-, амфирегулин, гепаринсвязывающий EGF (HB-EGF) и бетацеллулин. Как предполагают, EGF и TGF- являются основными эндогенными лигандами, которые вызывают EGFR-опосредованную стимуляцию, хотя, как показано, TGF- является более мощным в стимуляции ангиогенеза. Антагонисты EGFR также включают вещества, которые ингибируют димеризацию EGFR с другими субъединицами рецептора EGFR (т.е. гомодимеры EGFR) или гетеродимеризацию с другими рецепторами факторов роста (например, HER2). Антагонисты EGFR дополнительно включают биологические молекулы и малые молекулы, такие как синтетические ингибиторы киназ, которые действуют непосредственно на цитоплазматический домен EGFR для ингибирования EGFR-опосредованной передачи сигнала. Эрбитукс (цетуксимаб; С225) представляет собой пример антитела-антагониста EGFR, которое связывается с EGFR и блокирует связывание лиганда. Эрбитукс представляет собой химерное антитело IgG1, обладающее вариабельными доменами мыши из М225 (см., например, WO 96/40210) и константными доменами человека. Антитело анти-EGFR человека, обозначенное 11F8, описано в Zhu (WO 2005/090407). Другие антитела анти-EGFR включают EMD 72000 (матузумаб), вектибикс (панитумумаб; ABX-EGF), TheraCIM (нимотузумаб) и Hu-Max-EGFR (залутумумаб). Примером низкомолекулярного антагониста EGFR является ИРЕССА (ZD1939), который представляет собой производное хинозалина, функционирующее в качестве АТР-миметика для ингибирования EGFR. См. патент США № 5616582 (Zeneca Limited). Другим примером низкомолекулярного антагониста EGFR является ТАРЦЕВА (OSI-774), который представляет собой производное 4-(замещенный фениламино)хиноксалина [6,7-бис-(2-метоксиэтокси)хиназолин-4-ил]-(3-этинилфенил)амингидрохлорид] в качестве ингибитора EGFR. См. WO 96/30347 (Pfizer Inc.); Moyer et al., Cancer Res., 57:4838-48 (1997); Pollack et al., J. Pharmacol., 291:739-48 (1999). ТАРЦЕВА может функционировать путем ингибирования фосфорилирования EGFR и расположенных ниже него киназных путей передачи сигнала PI3/Akt и MAP (митоген-активированный белок), вызывая р27-опосредованную остановку клеточного цикла. См. Hidalgo et al., Abstract 281, представленный на 37th Annual Meeting of ASCO, San Francisco, CA, 12-15 May 2001. Хотя антагонисты можно вводить по отдельности, в определенных случаях желательно сочетать функции двух антагонистов в одной молекуле, такой как биспецифическое антитело или двойной ингибитор. Биспецифические антитела можно сконструировать так, чтобы сочетать специфичность в отношении IGF-IR со специфичностью в отношении другого RTK или другой молекулы на поверхности клетки. Особенно интересны сочетания специфичности в отношении IGF-IR со специфичностью в отношении EGFR или специфичностью в отношении HER2. Пример биспецифического антитела, которое связывается с IGF-IR и EGFR, представлен в Zhu (WO 2006/020258). Сходным образом, доступны малые молекулы, которые ингибируют IGF-IR и второй клеточный компонент, или можно осуществлять скрининг с их участием. Например, как упомянуто выше, INSM-18 (Insmed/University of California San Franscisco) ингибирует IGF-IR и HER2/neu. - 11 - 017265 Другой аспект настоящего изобретения относится к фармацевтическим композициям, содержащим антагонисты по настоящему изобретению или их фармацевтически приемлемую соль, гидрат или пролекарство, в сочетании с фармацевтически приемлемым носителем. Такие композиции могут являться отдельными композициями антагониста IGF-IR и средства ADT или одной композицией, содержащей и то и другое. Композиции по настоящему изобретению могут находиться в твердой или жидкой форме, в растворе или в суспензии. Способы введения включают, например, пероральное, парентеральное (внутривенное, внутрибрюшинное, подкожное или внутримышечное), местное, трансдермальное и путем ингаляции. Для перорального введения антагонист IGF-IR можно вводить, например, в жидкой форме с инертным разбавителем или усваиваемым носителем или включать в состав твердой лекарственной формы. Примеры пероральных жидких и твердых лекарственных форм включают, например, растворы, суспензии, сиропы, эмульсии, таблетки, пастилки, капсулы (включая мягкие желатиновые капсулы) и т.п. Пероральные лекарственные формы можно составлять в виде продуктов с замедленным высвобождением с использованием, например, покрытия для задержки распада или для контроля диффузии активного соединения. Там, где это необходимо, композиции могут также содержать солюбилизирующее средство. Примеры лекарственных форм для инъекций включают стерильные жидкости для инъекций, включая, например, растворы, эмульсии и суспензии. Лекарственные формы для инъекций включают твердые вещества, такие как стерильные порошки, которые заново разводят, растворяют или суспендируют в жидкости перед инъекцией. Стерильные растворы для инъекций получают внесением антагониста EGFIR и/или средства ADT в требуемом количестве в соответствующем растворителе с различными другими перечисленными выше ингредиентами, при необходимости с последующей стерилизацией фильтрованием. Носители, как правило, включают, например, стерильную воду, солевой раствор, инъецируемые органические сложные эфиры, арахисовое масло, растительное масло и т.п. Забуферивающие средства, консерванты и т.п. можно вносить в пригодных для введения формах. Стерильные композиции можно получать нагреванием, облучением, микрофильтрованием и/или добавлением различных антибактериальных и противогрибковых средств, таких как, например, парабены, хлорбутанол, фенол, сорбиновая кислота, тимерозал и т.п. Для местного введения антагонисты IGF-IR и средства ADT по настоящему изобретению можно вводить по отдельности или совместно, например, в форме гелей, кремов, или мазей, или лекарственных веществ в вязком носителе. Типичные носители для такого нанесения включают гидрофобные или гидрофильные основания, масляные или спиртовые жидкости и сухие порошки. Антагонисты IGF-IR и средства ADT можно также вносить в гель или матричную основу для нанесения при помощи лейкопластыря, необязательно, обеспечивая контролируемое высвобождение соединения через трансдермальный барьер. Антагонисты IGF-IR и средства ADT можно также составлять с помощью известных способов для ректального введения. Для введения путем ингаляции антагонисты IGF-IR и средства ADT по настоящему изобретению можно растворять или суспендировать в подходящем носителе или адсорбировать на нем для использования в распылительном, аэрозольном ингаляторе или ингаляторе с использованием сухих порошков. Подходящие дозировки может определять врач или квалифицированный медицинский специалист, и они зависят от факторов, таких как природа подлежащего лечению заболевания, способ введения, продолжительность лечения и состояние пациента. Антагонисты IGF-IR и средства ADT можно вводить так часто, как это необходимо для того, чтобы получить желаемый терапевтический эффект. Частота введения будет зависеть, например, от природы используемой лекарственной формы. Специалист в данной области может понимать, что дозировки и частота лечения зависят от переносимости конкретным пациентом и от фармакологических и фармакокинетических свойств используемого блокирующего или ингибирующего средства. В идеальном случае, желательно достижение насыщаемой фармакокинетики для используемого средства. Ударная доза для антитела анти-IGF-IR может варьировать, например, от приблизительно 10 до приблизительно 1000 мг/м2, предпочтительно от приблизительно 200 до приблизительно 400 мг/м2. За этим может следовать несколько дополнительных ежесуточных или еженедельных дозировок, варьирующих, например, от приблизительно 200 до приблизительно 400 мг/м2. Примерная дозировка антитела IGF-IR составляет 400 мг/м2 для ударной и 250 мг/м2 для еженедельной инфузии (для перерасчетов между мг/кг и мг/м2 для человека и других млекопитающих, см. Freireich, E.J. et al. , 1966, Cancer Chemother. Rep. 50:219-44). Пациента обследуют на побочные эффекты, и лечение останавливают, если такие побочные эффекты являются серьезными. Эффективные дозировки средств ADT хорошо известны в данной области. Специалист в данной области может также знать, как вести наблюдение за развитием лечения, чтобы определить эффективную дозу. Для рака предстательной железы один из таких способов представляет собой мониторирование уровней PSA. Другой способ состоит в мониторировании кислой фосфатазы предстательной железы (РАР). Другие способы мониторирования случаев рака предстательной железы включают ультразвук, компьютерную томографию (СТ), магнитно-резонансную томографию (MRI) и т.п. Образцы тканей можно исследовать на экспрессию и клеточное распределение AR, а также экспрес- 12 - 017265 сию сурвивина и/или TUBB. В некоторых вариантах осуществления изобретения лечение, сочетающее введение антагонистов IGF-IR с ADT, можно применять с одним или несколькими противоопухолевыми средствами. Например, как указано выше, ADT часто применяют в качестве неоадъюванта радиационной терапии опухолей предстательной железы. Если противоопухолевым средством является облучение, то источник облучения для подлежащего лечению пациента может являться или внешним (радиационная терапия от внешнего источника - EBRT), или внутренним (брахитерапия - ВТ). Противоопухолевое средство может представлять собой алкилирующее средство или антиметаболит. Примеры алкилирующих средств включают в качестве неограничивающих примеров цисплатин, циклофосфамид, мелфалан и дакарбазин. Примеры антиметаболитов включают в качестве неограничивающих примеров доксорубицин, даунорубицин и паклитаксел, гемцитабин. Пригодные противоопухолевые средства также включают ингибиторы митоза, такие как таксаны доцетаксел и паклитаксил. Ингибиторы топоизомеразы представляют собой другой класс противоопухолевых средств, которые можно применять в сочетании с антителами по изобретению. Они включают ингибиторы топоизомеразы I или топоизомеразы II. Ингибиторы топоизомеразы I включают иринотекан (СРТ-11), аминокамптотецин, камптотецин, DX-8951f, топотекан. Ингибиторы топоизомеразы II включают этопозид (VP-16) и тенипозид (VM-26). В качестве противоопухолевых средств в настоящее время оценивают другие вещества в отношении активности и эффективности в ингибировании топоизомеразы. В предпочтительном варианте осуществления ингибитор топоизомеразы представляет собой иринотекан (CPT-I1). По всему описанию изобретения цитируются различные публикации, тексты ссылок, руководства, технические пособия, патенты и патентные заявки. Идеи и описания этих публикаций, патентов, патентных заявок и других документов приведены в настоящем изобретении в качестве ссылки в полном объеме для более полного описания состояния области, к которой относится настоящее изобретение. Следует понимать и ожидать, что специалист в данной области может осуществлять вариации описанных здесь принципов изобретения, и подразумевается, что такие модификации должны входить в объем настоящего изобретения. Следующие примеры дополнительно иллюстрируют изобретение, но не следует полагать, что они каким-либо образом ограничивают объем изобретения. Подробные описания общепринятых способов, таких как способы, используемые в конструировании векторов и плазмид, и экспрессии антител и фрагментов антител, можно получить из многочисленных публикаций, включая Sambrook, J. et al. (1989), Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory Press; Coligan, J. et al. (1994), Current Protocols in Immunology, Wiley & Sons, Incorporated; Enna, S.J. et al. (1991), Current Protocols in Pharmacology, Wiley & Sons; Bonifacino, J.S. et al. (1999), Current Protocols in Cell Biology, Wiley & Sons. Все упомянутые здесь ссылки приведены в полном объеме. Примеры Антагонист IGF-IR ингибирует возобновление роста опухоли после ADT. Разработана преклиническая модель с кастрацией при возникновении рецидива рака предстательной железы после кастрации для тестирования эффективности ингибирования обусловленной IGF-IR передачи сигнала с использованием моноклонального антитела к IGF-IR человека (IMC-A12). Для исследования ксенотрансплантат LuCaP 35, андрогенчувствительной линии клеток рака предстательной железы человека, имплантировали подкожно в паховую область самцов мышей SCID. LuCaP 35 может переходить в андрогеннезависимое состояние, и ее можно использовать для оценки молекулярных изменений, связанных с этим процессом. Вначале происходит снижение уровней PSA и уменьшение объема опухоли, но после периода в 60-120 суток наблюдают возобновление роста опухолей. LuCaP 35 обладает метастатическим потенциалом и вызывает смешанные повреждения костей. LuCaP 35, растущая у интактных самцов мышей, является андрогенчувствительной и реагирует на андрогенную абляцию способом, который обычно наблюдают у пациентов. Клетки LuCaP 35 имплантировали подкожно в паховую область самцов мышей SCID. Когда опухоли достигали объема приблизительно 400 мм3, мышей кастрировали и разделяли на три группы из 20 животных в каждой. В контрольной группе 1 производили только кастрацию, в группе 2 производили кастрацию и вводили IMC-A12 внутрибрюшинно три раза в неделю в течение 14 суток, начиная с 7 суток после кастрации, и в группе 3 вводили IMC-A12 в течение 14 суток, начиная с 14 суток после кастрации. После 14 суток введения IMC-A12 не осуществляли никакого лечения. Расчет времени введения А12 в течение 2 недель, начиная с 1 или 2 недель после кастрации, производили на основе опубликованных данных для линии клеток LuCaP 35, которые указывают, что максимальная степень индуцируемого кастрацией апоптоза происходит в течение четырех суток после кастрации (Corey, E. et al., 2003, Prostate 99:392-401). Так как ингибирование обусловленной IGF-IR передачи сигнала может вызывать остановку клеточного цикла и предотвращать вступление клеток в апоптоз, было решено начинать введение А12, когда происходил "полный" апоптоз после кастрации (Corey et al., 2003; Tennant, M. et al., 2003, Prostate, 56:115-22). - 13 - 017265 Образцы крови собирали еженедельно из глазничного синуса. Сыворотку отделяли и уровни PSA определяли с использованием IMx Total PSA Assay (Abbott Laboratories, Abbott Park, IL). Опухоли измеряли два раза в неделю и объем опухоли оценивали по формуле: объем = L W2/2. Мышей умерщвляли при достижении опухолями размера 1000 мм3 или тогда, когда потеря массы животными превышала 20% первоначальной массы тела. BrdU вводили i.p. мышам за 1 ч перед тем, как животных умерщвляли, для определения in vivo степени пролиферации опухолевых клеток. После кастрации вначале происходила остановка роста опухолей у всех мышей (фиг. 1). У мышей, которым вводили IMC-А12, объем опухоли снижался в течение периода исследования, и у них отсутствовали случаи связанной с опухолями смерти. В необработанной группе увеличение среднего объема опухоли становилось выраженным к 5 неделе, при этом связанные с опухолями случаи смерти (когда животных умерщвляли) начинались на четвертую неделю и продолжались на протяжении исследования. Следует обратить внимание, что график среднего объема опухолей искусственно занижался для мышей, которые не получали IMC-A12, так как с каждым случаем смерти происходило удаление большой опухоли из усредненного множества опухолей. Уровни PSA отслеживали у мышей с ксенотрансплантатом LuCaP 35. Все мыши вначале реагировали на гормональную абляцию, и сходное снижение уровней PSA наблюдали в первую неделю после кастрации (фиг. 2). У мышей, которым производили только кастрацию, после начального снижения уровни PSA затем увеличивались в течение периода исследования, начиная приблизительно со второй недели. В то же время уровни PSA у кастрированных мышей, которым вводили IMC-A12, не увеличивались, но оставались вблизи нулевой линии. Это исследование показывает, что блокирование обусловленной IGF-IR передачи сигнала и экспрессии IGF-IR после кастрации при помощи антитела к IGF-IR, IMC-A12 приводит к значительно большему снижению объема опухоли по сравнению только с кастрацией, р<0,001 и значительно увеличивает время до возобновления AI-роста опухоли, который определяли по объему опухоли и увеличению уровня PSA, р<0,001. У контрольных животных, которым производили только кастрацию, рост опухоли прекращался приблизительно на четыре недели, но затем возобновлялся. Среди животных, которым производили только кастрацию, более половины было умерщвлено по причине роста опухоли к 9 неделям после кастрации, и большая часть животных была умерщвлена к концу 16 недель. В то же время, все животные, которые получали IMC-A12, оставались живыми после 16 недель. Представленные результаты in vivo говорят об эффективности ингибирования обусловленной IGFIR передачи сигнала. Примечательно, что антагонист IGF-IR вводили в течение 14 суток, и затем введение прекращали. В отдельном исследовании, в котором А12 вводили сходным образом, некоторое возобновление опухолевого роста наблюдали на поздней стадии исследования после введения А12. Двое из 40 животных групп 2 и 3 было умерщвлено в конце исследования по причине увеличения объема опухоли. Поддерживающие дозы антагониста IGF-IR, возможно, приведут к неопределенно долгому увеличению времени до возобновления опухолевого роста. Для исследования того, существует ли взаимосвязь между уменьшением объема опухоли у обработанных А12 опухолей и транслокацией AR, осуществляли иммуногистохимический анализ AR для опухолей из каждой трех групп, как представлено на фиг. 5. Показатель окрашивания ядер на AR определяли для 100 ядер из каждой опухоли. Ядра подсчитывались вслепую двумя индивидуумами, и среднее значение двух показателей рассматривали как показатель для данной ткани. Существует значимая положительная корреляция между объемом опухоли и интенсивностью ядерного AR, r=0,66, р0,01. Антагонист IGF-IR ингибирует транслокацию AR. Оценивали эффект стимуляции и ингибирования IGF-IR на локализацию рецептора андрогенов. Клетки LuCaP 35 культивировали в присутствии или в отсутствие стимуляции IGF-I, в присутствии или в отсутствие IMC-A12 (фиг. 3). Цитоплазматические и ядерные экстракты получали из обработанных клеток и оценивали с помощью PAGE. Уровень ERK использовали для выравнивания нагрузки на дорожки. В клетках, стимулированных IGF-I, IMC-A12 вызывал уменьшение доли наблюдаемого в ядре рецептора андрогенов. Транслокацию рецептора андрогенов также оценивали иммуногистохимией (фиг. 4). Опухоли из ксенотрансплантата LuCaP 35 (AD) пролиферировали в интактных самцах, и опухоли из ксенотрансплантата LuCaP 35V (AI) пролиферировали в кастрированных мышах. Тестируемым мышам вводили IMC-A12. Получали серийные срезы опухолей и окрашивали AR-специфическими антителами. У интактных контрольных мышей в андрогензависимых тканях LuCaP 35 AR локализовался преимущественно в ядре. В ткани тестируемых животных, которым вводили IMC-A12, окрашивание на AR наблюдали в цитоплазме. У кастрированных контрольных мышей в андрогеннезависимых клетках LuCaP 35v AR распределялся между ядром и цитоплазмой. В тканях тестируемых животных, которым вводили IMC-A12, окрашивание на AR наблюдали преимущественно в цитоплазме. - 14 - 017265 В сходном эксперименте локализацию AR исследовали флуоресцентной микроскопией в культуре тканей. Обработка 10-8 М DHT вызывала значительное перераспределение AR из цитоплазмы в ядро. Обработка только с помощью IGF-I вызывала частичное перераспределение AR в ядро, и IMC-A12 полностью обращал этот эффект. Антагонист IGF-IR ингибирует AR-зависимую экспрессию генов. Сурвивин, который является ингибитором апоптоза, обладает сильной экспрессией в нескольких линиях клеток рака предстательной железы человека. В линиях клеток с интактными рецепторами андрогенов андрогенная стимуляция посредством DHT усиливает экспрессию сурвивина. Также наблюдали, что экспрессия сурвивина опосредована Akt, так как передача сигнала от Akt, индуцированная IGF, усиливает экспрессию сурвивина даже в AR-негативных линиях клеток. Эксперимент с генным чипом для детектирования дифференциальной экспрессии сурвивина указывает на то, что экспрессия сурвивина снижается при лечении с помощью IMC-A12. Специальные микрочипы с кДНК конструировали, как описано ранее [ref] с использованием клонов, полученных из Prostate Expression Data Base (PEDB) , хранилища последовательностей для данных по экспрессирующимся маркерным последовательностям (EST) предстательной железы человека, находящихся в публичном доступе (Nelson, P.S. et al., 2002, Nucl. Acids Res. 30:218-20). Способы мечения с помощью флуоресцентных красителей Су3 и Су5, гибридизацию с панелям микрочипов и обработку чипов осуществляли, как описано (Tusher, V. et al., 2001, Proc. Natl. Acad. Sci. U.S.A. 98:5116-21). Отбирали по три опухоли из каждой экспериментальной группы. Для получения стандарта РНК для сравнения для использования в микрочипах с кДНК равные количества суммарной РНК выделяли и отбирали из линий клеток LNCaP, DU145, РС3 и CWR22rV1 (American Type Culture Collection, Manassas, VA), выращенных в логарифмической фазе в среде RPMI-1640 без красителя с добавлением 10% эмбриональной телячьей сыворотки (FBS; Life Technologies, Rockville, MD). Суммарную РНК выделяли из отобранных опухолей и линий клеток с использованием Trizol (Invitrogen, SanDiego, CA). мРНК амплифицировали в одном цикле с использованием набора для амплификации Ambion MessageAmp II (Ambion Inc, Austin, TX) и качество и количество образца оценивали с помощью агарозного гельэлектрофореза и поглощения при А260. Зонды для гибридизации метили и контроль качества для экспериментов с использованием чипов осуществляли, как описано ранее (Tusher, V. et al., 2001). Различия в экспрессии генов, связанные с обработанными группами, определяли с использованием процедуры SAM (Chu, G., Narasimhan, В., Tibshirani, R. & Tusher, V., 2002, Significance analysis of microarrays (sam) software, Stanford University) с рассматриваемой в качестве значимой (37) частотой ложных обнаружений (FDR) 10%. Сходства между образцами оценивали посредством неконтролируемой, иерархической кластеризации генов и образцов с использованием программного обеспечения Cluster 3.0 (de Hoon et al., 2004, Bioinformatics 20:1453-4) и просматривали с помощью Tree View (Page, R.D., 1996, Comput. Appl. Biosci. 12:357-8). Сурвивин и TUBB также оценивали с помощью PCR с использованием ранее описанных праймеров и способов (Wu, J. et al., 2006, Clin. Cancer Res. 12:6153-60). Очищали стандартный PCR-фрагмент кДНКмишени. Серию разведений стандартов от 10 нг/мкл до 10-3 пг/мкл использовали для RT-PCR в реальном времени для получения калибровочных кривых. 1 мкг суммарной РНК из каждой группы отобранных опухолей использовали для синтеза первой цепи кДНК с использованием Superscript First Strand Synthesis System (Invitrogen). RT-PCR в реальном времени осуществляли в 20 мкл реакционной смеси, состоящей из 1 мкл первой цепи кДНК, набора специфических праймеров и Lightcycler FastStart DNA Master Plus SYBR Green, с использованием Roche Lightcycler согласно инструкции производителя (Roche, Nutley, NJ). Продукты RT-PCR подвергали анализу кривой плавления посредством программного обеспечения Lightcycler v3.5. Размеры ампликонов подтверждали с помощью агарозного гель-электрофореза. Каждый образец анализировали в двух параллелях. Кастрация в сочетании с антагонистом IGF-IR связана со снижением экспрессии гена AR до тех пор, пока не произойдет рецидивирование опухоли. Образцы РНК из опухолей, полученные в каждой группе во временные рамки, указанные в табл. 2, анализировали на микрочипах с кДНК. Для группы 1 (только кастрация) не обнаружены гены, которые значительно изменены между периодами времени, при тестировании с помощью двухвыборочного t-критерия в SAM (добротность 100%). Кроме того, в неконтролируемой, иерархической кластеризации известных регулируемых андрогенами генов не разделялось два периода времени. Это может быть не удивительно, так как многие из животных в этой группе обладали восстановлением уровня PSA и увеличением показателей AR в ядре по сравнению с группами 2 и 3 к 40 суткам. В то же время имели место значительные изменения экспрессии генов между двумя периодами времени в обработанных А12 опухолях. Из 3170 уникальных генов на чипе с достаточным количеством данных для тестирования 21 подвергался стимуляции (включая многие регулируемые андрогенами) и 41 подвергался подавлению с 10% добротности в позднем периоде времени, когда опухоли начинали рецидивировать, по сравнению с ранним периодом времени (фиг. 6) Более того, посредством неконтролируемой, иерархической кластеризации известных регулируемых андрогенами генов отчетливо выделяются обработанные с помощью А12 два периода времени в двух отдельных кластерах. Эти - 15 - 017265 данные указывают на то, что экспрессия AR в ядре связана с транскрипционной активностью AR и прогрессированием рака предстательной железы посредством активации AR. Таблица 2 Чипы с кДНК в каждый момент времени Экспрессия сурвивина и -тубулина значительно снижалась посредством антагониста IGF-IR. В исследованиях с помощью микрочипов определили, что экспрессия сурвивина снижалась в опухолях, обработанных антителами А12. Как представлено на фиг. 7А, с помощью Qt-RT PCR с РНК, выделенной из опухолей, показана значимая положительная корреляция между количеством копий сурвивина и объемом опухоли, r=0,66, р0,01. Второй ген, для которого недавно обнаружено участие в индуцированном IGF-IR образовании опухолей, представляет собой -тубулин, TUBB (O'Connor, R., 2003, Horm. Metab. Res. 35:771-7; Geller, J. et al., 1984, J. Urol. 132:693-700). Показано, что количество TUBB снижено в микрочипах, и, как представлено на фиг. 7В, показано, что в образцах опухолей положительно коррелирует с объемом опухоли, r=0,59, р0,01, и значительно снижено в группах 2 и 3 по сравнению с группой 1. Третий ген, количество которого не отличалось с течением времени на микрочипах в группе 1, но снижалось в двух ранних периодах времени в группе животных 2 и 3, представлял собой PSA. Изменение экспрессии PSA подтверждено сходным профилем уровней PSA в сыворотке. Пролиферация и апоптоз. Апоптоз определяли анализом мечения концевого разрыва с помощью терминальной дезоксинуклеотидилтрансферазы (TUNEL) и окрашиванием пропидием (PI) с использованием набора Apop-Direct (BD BioScience), как описано ранее (Wu, J.D. et al., 2005, Clin. Cancer Res., 11:3065-74). В кратком изложении, 1106 клеток из клеточной суспензии фиксировали 10% формалином в нейтральном буфере (NBF) с последующей обработкой 70% этиловым спиртом при -20°С в течение 30 мин. После нескольких промывок клетки подвергали пермеабилизации 0,1% Тритоном Х-100 и инкубировали с dUTP, конъюгированным с FITC, и ферментом терминальной дезоксинуклеотидилтрансферазой (TdT) при 37°С в течение 1 ч, с последующей инкубацией с буфером с PI/РНКазой (100 мкг/мл PI, 50 мкг/мл РНКазы) при комнатной температуре в течение 60 мин. Образцы анализировали проточной цитофлуориметрией с использованием BD FACscan. Данные анализировали с помощью программного обеспечения CellQuestPRO. Апоптоз также определяли с использованием анализа TUNEL в фиксированной формалином ткани с использованием набора Apop-Tag (Millipore Co, MA) согласно инструкциям производителя. Апоптотические клетки определяли на 300 клеток на образец ткани. Как представлено в табл. 3, пролиферация оказалась значительно выше в группе 1 опухолей по сравнению с группой 2 и 3, р0,01. В то же время апоптоз при определении окрашиванием с помощью TUNEL оказался выше в группе 1 по сравнению с группами 2 и 3, табл. 3. Таблица 3 Апоптоз и поглощение BRDU **р<0,001 по сравнению с группой с кастрацией. - 16 - 017265 Список последовательностей - 17 - 017265 - 18 - 017265 - 19 - 017265 - 20 - 017265 - 21 - 017265 - 22 - 017265 - 23 - 017265 - 24 - 017265 - 25 - 017265 - 26 - 017265 - 27 - 017265 - 28 - 017265 - 29 - 017265 - 30 - 017265 - 31 - 017265 - 32 - 017265 - 33 - 017265 - 34 - 017265 - 35 - 017265 - 36 - 017265 - 37 - 017265 ФОРМУЛА ИЗОБРЕТЕНИЯ 1. Применение антитела IMC-A12, которое является ингибитором IGF-IR, для отдельного, совместного или последовательного введения с андрогенной депривацией для лечения рака предстательной железы. 2. Применение антитела IMC-A12, которое является ингибитором IGF-IR, для отдельного, совместного или последовательного введения с андрогенной депривацией для ингибирования или предотвращения перехода андрогензависимого рака предстательной железы в андрогеннезависимый рак предстательной железы. 3. Применение по п.1 или 2, где андрогенная депривация включает введение аналога рилизингфактора лютеинизирующего гормона (LHRH). 4. Применение по п.1 или 2, где андрогенная депривация включает проведение лечения антиандрогенами. 5. Применение по п.1 или 2, где андрогенная депривация включает введение ингибитора андрогенов надпочечников. 6. Применение по п.1 или 2, где андрогенная депривация включает орхиэктомию. 7. Применение по п.1 или 2, где дополнительно используют антагонист Akt в сочетании с IMC-A12. 8. Применение по п.7, где антагонист Akt является антагонистом EGFR. 9. Применение по п.1 или 2, где дополнительно используют противоопухолевое средство в сочетании с IMC-A12. 10. Применение по п.9, где противоопухолевым средством является облучение. 11. Способ лечения рака предстательной железы, включающий введение IMC-A12 и андрогенную депривацию, которые начинают приблизительно в одно и то же время. 12. Способ лечения рака предстательной железы, включающий введение IMC-A12 и андрогенную депривацию, в котором IMC-A12 вводят после депривации андрогенов и перед тем, как рак предстательной железы переходит от андрогензависимого рака к андрогеннезависимому раку. Фиг. 1 - 38 - 017265 Фиг. 2 Фиг. 3 Фиг. 4 - 39 - 017265 Фиг. 5 Фиг. 6 - 40 - 017265 Фиг. 7 Евразийская патентная организация, ЕАПВ Россия, 109012, Москва, Малый Черкасский пер., 2 - 41 -