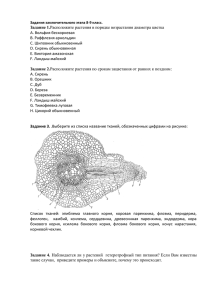
АНАТОМИЯ РАСТЕНИЙ
реклама

В.Б. СКУПЧЕНКО АНАТОМИЯ РАСТЕНИЙ Учебное пособие САНКТ-ПЕТЕРБУРГ 2004 Министерство образования и науки Российской Федерации САНКТ-ПЕТЕРБУРГСКАЯ ГОСУДАРСТВЕННАЯ ЛЕСОТЕХНИЧЕСКАЯ АКАДЕМИЯ В.Б. СКУПЧЕНКО, доктор биологических наук АНАТОМИЯ РАСТЕНИЙ Учебное пособие для студентов специальностей «Лесное хозяйство» и «Садово-парковое и ландшафтное строительство» направления «Лесное хозяйство и ландшафтное строительство» Допущено Учебно-методическим объединением по образованию в области лесного дела в качестве учебного пособия для студентов высших учебных заведений специальностей 260400 «Лесное хозяйство» и 260500 «Садово-парковое и ландшафтное строительство» направления 656200 «Лесное хозяйство и ландшафтное строительство» Санкт-Петербург 2004 1 Рассмотрено и рекомендовано к изданию учебно-методической комиссией лесохозяйственного факультета Санкт-Петербургской государственной лесотехнической академии 16 октября 2003 г. Рецензенты: кафедра ботаники биолого-почвенного факультета Санкт-Петербургского государственного университета (зав. кафедрой, профессор А.А. Паутов), С.В. Загирова, доктор биологических наук (Институт биологии Коми НЦ УРО РАН) УДК 630 Скупченко В.Б. Анатомия растений: Учебное пособие для вузов. СПб.: СПбГЛТА, 2004. 180 с. Представлена характеристика строения и функций клеток, тканей и вегетативных органов семенных растений. Дано описание структуры древесины некоторых голосеменных и покрытосеменных растений. Приведены сведения о микроскопической технике и принципах подготовки объектов к анатомическому изучению. Для студентов лесотехнических вузов. Ил. 108. Библ. 19 назв. Темплан 2004 г. Изд. № 20. ISBN 5–9239–0016–5 2 © Санкт-Петербургская государственная лесотехническая академия (СПбГЛТА), 2004 ПРЕДИСЛОВИЕ Настоящее пособие составлено в соответствии с содержанием раздела Анатомия растений программы дисциплины «Ботаника» высших учебных заведений для специальности 260400 «Лесное и лесопарковое хозяйство», которая утверждена Государственным комитетом Российской федерации по высшему образованию в 1995 г. Анатомия растений представляет собой важный раздел ботаники и служит основой для освоения студентами ряда биологических дисциплин, включающих физиологию растений, общую экологию, фитопатологию, древесиноведение и др. В соответствии с этим, в данном пособии уделяется внимание структурной и функциональной характеристике основных уровней организации растительного организма – субклеточного, клеточного, тканевого, структуры органов и системы их взаимосвязи в целом растении. Важное внимание уделяется вопросам происхождения и развития клеток и тканей из соответствующих образовательных тканей и выполнению ими определенных функций. Наряду с описанием основных общих понятий анатомии растений, данное пособие содержит также анатомическую характеристику ряда растительных объектов, сведения об устройстве микроскопов, методику их применения и способы настройки. Приведены общие принципы подготовки объектов для их изучения при помощи микроскопов. Иллюстрации составлены из оригинальных анатомических рисунков, выполненных автором пособия, и рисунков, заимствованных из ряда отечественных и зарубежных руководств и научных изданий следующих авторов: Барыкина Р.П. и др.(1979); Грин Н. и др. (1990); Еремин В.М. (1978); Лотова Л.И., Тимонин А.К. (1989); Палладина В.И. (1895); Хржановский В.Г., Пономаренко С.Ф. (1979); Рейвн П. и др. (1990); Эсау К. (1969, 1980); Яковлев Г.П., Челомбитько В.А. (1990); Яценко-Хмелевский А.А. (1961); К.Н. Демченко, Н.П. Демченко (2001), а также из Атласа ультраструктуры растительных тканей (1980) и других источников. Автор сердечно благодарит профессора А.А. Паутова (СПбГУ), доктора биологических наук С. В. Загирову (Институт биологии Коми НЦ УрО РАН), профессора О.И. Полубояринова, а также сотрудников кафедры общей экологии, анатомии и физиологии растений ЛТА за ценные замечания и предложения, направленные на совершенствование рукописи. 3 Г л а в а 1. В В Е Д Е Н И Е Анатомия растений (греч. «анатоме» – рассекаю) – это раздел ботаники, занимающийся изучением внутреннего строения растений. Анатомия растений представляет собой часть более общей ботанической дисциплины – морфологии растений. Основной метод анатомии состоит в рассечении различных органов растений на тонкие слои и их изучение при помощи световых и электронных микроскопов. В зависимости от направления приложения сведений о внутренней структуре объектов, анатомия растений подразделяется на ряд следующих разделов. Описательная анатомия растений занимается изучением внутреннего строения зрелых органов в их окончательном, дефинитивном, состоянии, при этом не рассматривается вопрос о развитии этих структур. Анатомия развития, или онтогенетическая анатомия, занимается изучением данных о строении растений с точки зрения постепенного развития структур в онтогенезе. Эволюционная, или филогенетическая, анатомия растений занимается изучением преобразований признаков структуры в процессе эволюции – исторического развития. Сравнительная, или систематическая, анатомия растений занимается описанием диагностических или систематических признаков, характеризующих определенную систематическую категорию – таксон (вид, род, семейство и т.п.). Экологическая анатомия осуществляет исследования, направленные на раскрытие связей между строением растения и условиями его существования. Большое значение имеет анатомия растений в идентификации (определении) продуктов растительного происхождения. Например, при анализе сухих кормов для животных, некоторых пищевых продуктов, используемых в сухом виде (чай, табак, различные пряности и т.д.). Микроскопический анализ применяется для установления подлинности лекарственного сырья на основе признаков анатомического строения растений в фармакогнозии. Довольно часто возникает необходимость определить растение по анатомическим признакам, которые сохраняются после длительного хранения или в ре4 зультате сложной механической обработки, например измельчении до порошка. С помощью анатомического метода решаются многие важные вопросы древесиноведения. Так, например, из древесины, имеющей различные качество и ценность, могут быть изготовлены разнообразные материалы и изделия. На основе анатомического анализа по микроскопическим признакам можно достаточно точно установить принадлежность ксилемы к определенной древесной породе. Для этого подготовлены специальные пособия, определители и атласы, описывающие строение деловых древесин, применяемых в строительстве и деревообрабатывающей промышленности. Анатомический контроль применяется на различных этапах производства бумаги. Анатомический метод изучения растительных остатков достаточно широко применяется в криминалистической практике. В палеоботанике – науке об ископаемых растениях, применяется анатомический метод для изучения древних представителей флоры по остаткам их органов, подвергнутых сложным процессам минерализации. Палеоботанические сведения широко применяются в геологии, например, при определении датировки геологических отложений в стратиграфии. В области изучения болезней растений важное место занимает анатомический метод исследования. С его помощью решается вопрос, каким образом инфицируются органы и ткани растений и как они реагируют на внедрение вредных организмов. Анатомия растений имеет большое значение при установлении разнообразных жизненных функций организмов, которые изучает физиология растений. Анатомический метод позволяет с высокой степенью точности выявить локализацию тех или иных процессов в растительном организме, а также места накопления, депонирования продуктов жизнедеятельности. Это становится возможным благодаря использованию методов гистохимии, люминесцентной микроскопии, наблюдения объектов в ультрафиолетовом свете и цитофотометрии. Особенно эффективна в этом отношении электронная микроскопия в сочетании с методами электронной гистохимии и морфометрическим количественным анализом ультрастуктур клетки. 5 КРАТКИЙ ОБЗОР АНАТОМИЧЕСКОЙ ТЕХНИКИ И МЕТОДОВ ПОДГОТОВКИ ОБЪЕКТОВ К ИССЛЕДОВАНИЮ Невооруженный глаз человека со средней остротой зрения в две угловые минуты, наблюдая хорошо освещенный предмет с расстояния наилучшего видения – 250 мм, может различать отдельные детали размером не менее 0,15 мм. Разрешающая способность обычного светового микроскопа составляет десятые доли микрометра (0,2–0,3 мкм) наблюдаемого объекта. Таким образом, полезное увеличение светового микроскопа может достигать более 1000 раз. В электронном просвечивающем микроскопе можно различать детали объекта минимальным размером до 0,1–0,2 нм, то есть в 1000 раз более мелкие, чем в световом, а максимальные увеличения могут достигать несколько сот тысяч раз. Световой микроскоп позволяет рассмотреть ткани, клетки и отдельные крупные клеточные структуры – ядро, ядрышки, цитоплазму, пластиды, вакуоли, клеточную оболочку и др. В электронном микроскопе имеется возможность подробно изучать тонкую, субклеточную структуру – оболочки клетки, органелл, биологических мембран, а также непосредственно наблюдать макромолекулярные образования нуклеиновых кислот, белков, полисахаридов и др. Анатомическое исследование растительного объекта может быть осуществлено после его предварительной подготовки, которая зависит от задачи исследования и характера последующего изучения при помощи различных типов микроскопов. Наиболее распространена просвечивающая микроскопия объектов, предусматривающая их рассмотрение в проходящем свете или потоке электронов. Для такого способа анатомического изучения объекты предварительно фиксируются этиловым спиртом, альдегидами или солями тяжелых металлов для сохранения прижизненного состояния клеточных структур и тканей. После чего фиксирующий раствор отмывается, и объект подготавливается к разрезанию (срезы толщиной 5–30 мкм для световой микроскопии и 0,02–0,06 мкм для электронной микроскопии). Основной формой изучения объектов в световом микроскопе служит светомикроскопический препарат. Он представляет собой основание в виде предметного стекла размером 2 х 25 х 75 мм, на котором размещается срез объекта в заключающей среде, твердой или жидкой, накрытый тонким покровным стеклом, размером 0,18 х 6 20 х 20 мм. Для световой микроскопии объекты традиционно послойно разрезаются на салазочных (рис. 1) или ротационных микротомах путем состругивания тонких срезов. Перед срезанием зафиксированные образцы обязательно пропитываются твердым средами – парафином или целлоидином по соответствующим гистологическим методикам. Пропитывание твердыми средами можно исключить в случае применения замораживающих или новейших вибрационных микротомов. Срезы затем монтируются на предметном стекле, окрашиваются красителями общего назначения – сафранином, метиленовым синим или кармином, либо гистохимическими красителями для выявления соответствующих веществ клетки. После этого срезы заливаются затвердевающей заключающей средой, например канадским бальзамом, и накрываются тонким покровным стеклом. Рассматривание образцов может быть выполнено в видимом свете при помощи обычного биологического или рабочего микроскопа, а также при помощи ультрафиолетового или люминесцентного микроскопов в ультрафиолетовом свете, либо при помощи инфракрасного микроскопа в инфракрасной области спектра. Кроме того, можно наблюдать двойное лучепреломление объектов (оболочек клеток, гранул крахмала, кристаллов) при помощи поляризационного микроскопа. Большой прогресс в исследовании и микрокиносъемке жизнедеятельности клеток и тканей достигнут с использованием метода фазового контраста, повышающего четкость изображения живых объектов без их дополнительного окрашивания. Изучение объектов в просвечивающем трансмиссионном электронном микроскопе выполняется на ультратонких срезах, помещенных на тонкие медные сетки диаметром 2,5-3,0 мм. При этом срезы удерживаются на поверхности сетки силами адгезии. Ультратонкие срезы получают на ультрамикротоме (после пропитывания объекта специальными эпоксидными смолами и их полимеризации по соответствующим методикам). В качестве ножа для такого микротома применяют трехгранную призму из стекла толщиной около 8 мм. Она получается путем раскола по диагонали стеклянного квадрата со стороной 25 мм на специальном станке. Около режущего ребра стеклянного ножа монтируется ванночка, в которую наливается дистиллированная вода. Полученные ультратонкие срезы, площадью до 1 мм2, располагаются на поверхности воды в ванночке в виде окрашенных интерферирующих пленок. Срезы оптимальной толщины около 40–50 нм интерферируют желтым золотистым цветом. Серии ультратонких 7 срезов, плавающие на поверхности воды в ванночке, монтируются на тщательно очищенные медные сетки. После соответствующих процедур контрастирования солями тяжелых металлов ультратонких срезов, расположенных на медной сетке, ее помещают в глубокий вакуум колонны микроскопа в узкий пучок электронов, ускоренных напряжением до 100000 В. Кроме просвечивающей микроскопии существует микроскопия в отраженном свете. Для световой отраженной микроскопии не требуется специальной подготовки объекта, и он может быть рассмотрен в естественном виде. Однако такая микроскопия обладает небольшой глубиной резкости, и она не позволяет рассматривать в фокусном состоянии структуры объекта, имеющие глубокий рельеф. Современные конфокальные световые микроскопы, используют для освещения узкий луч лазера, сканирующий по объекту, вызывая его люминесценцию. Свет люминесценции от каждой конкретной точки объекта направляется через узкую диафрагму на фотоэлектронный умножитель, преобразуется в электрический сигнал, который усиливается и после обработки компьютером направляется на определенную часть изображения на мониторе. Данные микроскопы обладают повышенной разрешающей способностью, возможностью трехмерной реконструкции объектов и морфометрической обработки изображений по соответствующим программам. Особый прогресс в изучении самого сложного рельефа объектов достигнут при помощи сканирующей электронной микроскопии, благодаря ее практически неограниченной глубине резкости. В сканирующем электронном микроскопе – стереоскане очень узкий луч электронов пробегает, сканирует последовательно по линиям весь наблюдаемый объект. Поверхность образца для предотвращения электризации предварительно напыляется в вакууме ровным и тонким слоем токопроводящего материала – золотом, углеродом и т.п. Торможение электронов поверхностью объекта вызывает ионизирующее излучение, которое улавливается специальным приемником – детектором. Усиленный и обработанный электронными устройствами сигнал детектора направляется на электронно-лучевую трубку или фотопластинку, где и получается очень рельефное объемное изображение объекта. 8 Рис. 1. Санный микротом с подъемным объектодержателем, движущимся по наклонной плоскости: 1 – станина; 2 – ножевые салазки с ножом; 3 – микрометрический винт; 4 – рамка с ограничителем; 5 – салазки с объектодержателем НАСТРОЙКА ПРОСВЕЧИВАЮЩЕГО СВЕТОВОГО МИКРОСКОПА Микроскоп устанавливается у левого глаза наблюдателя (рис. 2, А, Б). С правой стороны на столе помещается альбом для зарисовок. Рукояткой грубой фокусировки тубусодержатель приподнимается над предметным столиком. На оптическую ось устанавливается до щелчка, устойчивого фиксированного положения, объектив с 8-кратным увеличением. Рукояткой грубой фокусировки объектив приближается к предметному столику на расстояние 8–10 мм. Конденсор поднимается вверх до упора. Диафрагма конденсора открывается движением по часовой стрелке. Глядя в окуляр, движением зеркала улавливается источник света, который центрируется так, чтобы свет равномерно и ярко освещал все поле зрения. 9 А Рис. 2. Световые микроскопы: А – МБС-1, Б – Биолам: 1 – окуляр, 2 – тубус, 3 – тубусодержатель, 4 – рукоятка грубой фокусировки, 5 – рукоятка тонкой фокусировки, 6 – подставка, 7 – зеркало, 8 – конденсор с ирисовой диафрагмой, 9 – предметный столик, 10 – револьвер с объективами, 11 – стойка, 12 – оптическая головка, 13 – объектив, 14 – рукоятка переключения увеличения, 15 – бинокулярная насадка Микропрепарат помещается на предметный столик микроскопа таким образом, чтобы покровное стекло находилось сверху. Смотреть в микроскоп рекомендуется левым глазом. Переход с малого увеличения к большому, например с объективом 40х, выполняется в следующем порядке. Первоначально интересующий участок объекта помещается в центр поля зрения. Затем рукояткой грубой фокусировки тубус микроскопа слегка приподнимается на 1 – 2 мм. После этого на оптическую ось до устойчивого фиксированного положения устанавливается объектив 40х. Рукояткой грубой фокусировки объектив приближается к покровному стеклу микропрепарата примерно на 0,5 мм. Затем рукояткой тонкой фокусировки достигается четкое изображение объекта. Ряд объективов большого увеличения (40х, 60х, 70х, 90х) применяется с иммерсионной жидкостью, имеющей больший показатель преломления, чем воздух. Этой жидкостью заполняют пространство между первой линзой объектива и покровным стеклом микропрепарата, увеличивая числовую апертуру объектива и, следовательно, разрешающую способность микроскопа. Объективы, применяемые с водной иммерсией, имеют на оправе белый ободок, а с масляной иммерсией (кедровое масло) – черный. 10 КЛАССИФИКАЦИЯ ТКАНЕЙ РАСТЕНИЙ Ткани представляют собой системы клеток, сходные по строению, функциям и происхождению. Все высшие растения состоят из тканей. Ткань относится к простой в том случае, когда она образована морфологически и функционально одним типом клеток. Существуют также сложные комплексные ткани, состоящие из нескольких типов клеток. В зависимости от выполняемой функции ткани подразделяются на несколько групп: образовательные (меристемы), покровные, проводящие, механические, основные, выделительные (секреторные). Способность к делению – характерная черта клеток меристем. Все остальные ткани относятся к постоянным, они развиваются из меристем. В процессе дифференциации меристематические клетки, имеющие одинаковое происхождение, приобретают индивидуальные функциональные и морфологические отличия, характерные для постоянных тканей. Это сложный процесс, связанный с действием различных факторов, в том числе и ростовых веществ – гормонов. Дифференцированные клетки, имеющие живые протопласты с полным набором органелл, в определенных условиях, вторично приобретают способность к делению. РАЗВИТИЕ И ОБЩАЯ СТРУКТУРА РАСТИТЕЛЬНОГО ОРГАНИЗМА Тело высшего растения представляет собой спорофитную фазу жизненного цикла и начинает свое существование от одноклеточной зиготы – оплодотворенной яйцеклетки, из которой, путем организованных делений и роста клеток, развивается зародыш. Зародыш имеет относительно простое строение (рис. 3). Он содержит ось, от которой отходит одно или несколько листоподобных образований – семядолей. Выше семядолей ось переходит в полюс побега, который может быть представлен только апикальной мери11 стемой побега, либо зародышевым побегом, называемым эпикотилем или плюмулой. Ниже семядолей расположена стеблевая часть зародыша – гипокотиль. За гипокотилем следует корневой полюс, на котором располагается зачаток корня, представленный апикальной меристемой корня, либо зародышевым корешком. Рис. 3. Схема строения зародыша двудольного растения. I – поперечное сечение через апекс побега и основание семядолей; II – продольное сечение зародыша. 1 – семядоли, 2 – прокамбиальные пучки, 3 – апикальная меристема побега, 4 – гипокотиль, 5 – апикальная меристема корня, 6 – корневой чехлик Ткани зародыша остаются еще меристематическими, но по расположению и клеточным характеристикам начинают приобретать признаки сходства с тканями, которые формируются в развивающемся из зародыша проростке. Из наружного слоя клеток – протодермы, формируется первичная покровная ткань – эпидерма. Под нею расположены вакуолизированные клетки основной меристемы будущей коры. Вдоль всего зародыша по оси зачатка корня, гипокотиля и семядолей проходит система прокамбиальных пучков, из которых впоследствии развивается проводящая система растения, сложенная первичной флоэмой и первичной ксилемой. Апикальная меристема побега, после прорастания семени, начинает формировать меристематические структуры узлов, междоузлиев и зачатков листьев. В пазухах листьев формируются новые апексы побегов, из которых развиваются пазушные побеги. Таким образом, на главном стебле растения формируется система ветвей. Из апикальной меристемы корня зародыша развивается главный (первичный) корень. Боковые (вторичные) корни формируются из новых апикальных меристем, закладывающихся эндогенно на главном корне. В результате этого развивается система ветвления корней. У взрослых растений, главным образом, однодольных, формируются придаточные (адвентивные) 12 корни, которые развиваются на стебле, образуя мочковатую корневую систему, в виде щетки или кисти, сходных по длине и форме корней. У древесных растений апикальные меристемы побега и корня функционируют в продолжение многих лет, формируя ежегодно зачатки новых побегов и корней. Ранее сформированные побеги и корни, как правило, на второй год прекращают первичный продольный рост, но продолжают радиальный рост (по толщине) в течение многих лет, благодаря образованию длительно функционирующих вторичных латеральных меристем – камбия и феллогена. Камбий обеспечивает ежегодное формирование новых клеток проводящих тканей – вторичной флоэмы и вторичной ксилемы. Феллоген формирует вторичную покровную ткань – перидерму. Таким путем семенное растение осуществляет рост на вегетативной стадии жизни растения. За вегетативной стадией в жизни растения следует репродуктивная. Переход к репродуктивной стадии связан с эндогенным ритмом роста и действием внешних факторов – температурой, световым режимом и др. В продолжение ее вегетативный апекс побега превращается в репродуктивную (генеративную) апикальную меристему, из которой у цветковых растений развивается цветок или соцветие, а у голосеменных – микростробил или макростробил. В настоящее время еще относительно мало известно о факторах, управляющих активизацией деления клеток, ориентацией направления роста меристем, характером дифференциации и специализации клеток при переходе к выполнению определенных функций. Сведения об этих актуальных направлениях биологии растений могут быть существенно расширены с применением современных разработок в области анатомии и физиологии растений, биохимии, микроскопической техники, компьютерного морфометрического анализа изображений и др. Положение тканей в растительном организме можно рассмотреть на примере растения льна (рис. 4). На верхушке стебля находится апикальная меристема побега. Она формирует новые структуры стебля – узлы и междоузлия, а также листовые зачатки. Вблизи апикальной меристемы стебель 13 Рис. 4. Структура сосудистого растения. I – общий вид растения льна (Linum usitatissimum) в вегетативной стадии, II – верхушка побега с апикальной меристемой и развивающимися листьями, III – окончание корня с корневым чехликом и апикальной меристемой, IV – листовая пластинка, V – стебель первичной структуры, VI – стебель вторичной структуры, VII – корень вторичной структуры, VIII – корень первичной структуры (II–III – продольное сечение, IV–VIII – поперечное сечение). 1 – апекс побега, 2 – молодые листья, 3 – первичная кора, 4 – прокамбий, 5 – листовая щель, 6 – ризодерма, 7 – боковая жилка, 8 – центральный цилиндр, 9 – апекс корня, 10 – корневой чехлик, 11 – эпидерма, 12 – ассимиляционная паренхима (мезофилл), 13 – проводящие пучки, 14 – первичная ксилема, 15 – первичная флоэма, 16 – сердцевина, 17 – листовые основания, 18 – волокна первичной флоэмы, 19 – вторичная флоэма, 20 – камбий, 21 – вторичная ксилема, 22 – радиальные лучи, 23 – феллема (пробка), 24 – перицикл, 25 – эндодерма, 26 – пазушный побег 14 и листья еще продолжают продольный и объемный рост и имеют признаки первичного строения (рис. 4, V). По мере дифференциации ранее заложенных структур, расположенных в средней части побега, происходит остановка их первичного роста и прекращается продольное удлинение (рис. 4, VI). Наряду с этим, здесь в стебле развивается вторичная латеральная меристема, которая начинает продуцировать клетки проводящих тканей – вторичной ксилемы и вторичной флоэмы. Они увеличивают объем стебля и обусловливают продолжение его радиального роста. Листовые зачатки, по мере удаления от растущей верхушки побега, разрастаются в зрелые листья, способные к активному фотосинтезу, с развитой ассимиляционной паренхимой, разветвленной системой проводящих пучков (жилок) и активно функционирующими устьицами в эпидерме (рис. 4, IV). Разветвленная корневая система имеет множество окончаний. Каждое из них покрыто корневым чехликом (рис. 4, III). Под чехликом располагается апикальная меристема корня. Ее инициальные клетки формируют во внешнюю сторону новые слои клеток корневого чехлика, сменяющие постоянно слущивающиеся их наружные слои. Во внутреннюю сторону инициальные клетки осуществляют формирование новых слоев меристематических клеток, которые обеспечивают продольный рост корня и постоянное нарастание его тканей: ризодермы, коры и центрального цилиндра. В этой части корень имеет первичное строение. Выше зоны всасывания в корне начинают проявляться признаки вторичного роста – появляется камбий и феллоген (рис. 4, VII). Камбий продуцирует новые клетки вторичной ксилемы и вторичной флоэмы, а феллоген – пробки и феллодермы, относящиеся к элементам перидермы – вторичной покровной ткани. Это обусловливает вторичный радиальный рост корня. 15 Г л а в а 2. К Л Е Т К А Клетка – это основная структурная и функциональная единица живых организмов. Клетка устроена так, что может сохранять и переносить наследственную информацию, в результате чего ее развитие и развитие ее потомства происходит упорядоченным образом. Таким путем поддерживается целостность организма. В многоклеточном организме клетки связаны друг с другом и функционируют согласованно, как единая система. Термин клетка (лат. Cellule – полость) впервые применил в 1665 г. Роберт гук при описании срезов пробки. Клеточное строение организмов как универсальная форма организации всего живого обосновано клеточной теорией, сформулированной в 1839 г. Ботаником М.Я. Шлейденом и зоологом т. Шванном. Согласно современным представлениям клеточная теория исходит из единства расчлененности многоклеточного организма на клетки и его целостности, основанной на взаимодействии клеток. Значительный прогресс в изучении клетки наступил с 1946 г. Благодаря широкому внедрению в биологию электронного микроскопа, обладающего большей чем в 1000 раз разрешающей способностью по сравнению со световым микроскопом. С помощью электронного микроскопа была подробно изучена субмикроскопическая структура известных из световой микроскопии компонентов клетки – ядра, пластид, митохондрий клеточной оболочки, а также выявлен целый ряд новых структурных клеточных элементов, например эндоплазматическая сеть, рибосомы, элементы цитоскелета и др. Живое содержимое клетки – протопласт окружено снаружи плазматической мембраной – плазмалеммой, которая, в свою очередь, покрыта клеточной оболочкой. Большинство клеток растений, а также некоторых тканей животных образует твердую оболочку, представляющую собой продукт жизнедеятельности протопласта. Плазмалемма выполняет функции активного физиологического барьера между протопластом и окружающей средой. В клетке осуществляется синтез веществ, а также преобразование энергии запасных органических веществ в форме легко доступных макроэргических соединений, например АТФ. Эти соединения используются для поддержания разнообразных процессов жизнедеятельности клетки: ее роста и метаболизма, включающего реакции синтеза веществ (катаболизма) и их расщепления (анаболизма). По степени сложности существуют два основных типа клеток. Первый тип – прокариотические клетки. Они имеют простое морфологическое строение и не содержат оформленного ядра, а их вещество наследственности ДНК находится прямо в цитоплазме и не окружено мембранной оболочкой. Органелл в таких клетках мало, и они не имеют мембранных оболочек. К организмам, для которых характерны прокариотические клетки, относятся бактерии и сине-зеленые водоросли. 16 Клетки второго типа – эукариотические – крупнее, чем прокариотические, из них состоят все организмы, за исключением бактерий и сине-зеленых водорослей. Эукариотические клетки содержат множество ограниченных мембранами органелл и немембранных структур, выполняющих разные функции. Каждая растительная клетка (рис. 5) снаружи окружена оболочкой, основу структуры которой составляют полисахариды. Под оболочкой находится клеточная мембрана плазмалемма, окружающая живое содержимое клетки – протопласт. Он состоит из жидкой коллоидальной среды – цитозоля, называемой также основной цито- Рис. 5. Ультраструктурная организация растительной клетки: 1 – оболочка одной клетки, 2 – плазмодесма, 3 – срединная пластинка, 4 – оболочки двух смежных клеток, 5 – свободные рибосомы в цитоплазме, 6 – цистерна шероховатого эндоплазматического ретикулума, 7 – рибосомы на поверхности шероховатого эндоплазматического ретикулума, 8 – хлоропласты, 9 – две мембраны оболочки хлоропласта, 10 – тилакоиды граны, 11 – митохондрия, 12 – диктиосома – тельце гольджи, 13 – клеточный сок вакуоли, 14 – тонопласт, окружающий вакуоль, 15 – пузырек гольджи, 16 – цистерна диктиосомы, 17 – микрофиламенты, рассеянные по всей цитоплазме, 18 – микротрубочки, 19 – цитоплазма, 20 – плазмалемма – наружная клеточная мембрана, 21 – гладкий эндоплазматический ретикулум, 22 – эухроматин, 23 – гетерохроматин, 24 – ядрышко, 25 – ядерная пора, 26 – ядерная оболочка, состоящая из двух мембран 17 плазмой, в которую погружены ядро, с ядрышками, пластиды, митохондрии, диктиосомы, эндоплазматический ретикулум, рибосомы, вакуоли, микротельца, микротрубочки, микрофиламенты. Важная роль в клетке принадлежит мембранам. Запасные вещества и отбросы, образующиеся в процессе жизнедеятельности клетки, называются эргастическими веществами. В клетках присутствуют и другие структуры, роль которых окончательно не выяснена. В отличие от большинства животных клеток, клетки растений имеют следующие специфические структуры: 1) жесткую полисахаридную клеточную оболочку (в отличие от белковой у большинства животных); 2) плазмодесмы в порах клеточной оболочки, через которые осуществляется взаимосвязь протопластов соседних клеток; 3) крупные вакуоли, ограниченные мембраной тонопластом, – вместилища клеточного сока; 4) пластиды, к которым относятся хлоропласты, зеленые пластиды, осуществляющие процесс фотосинтеза органических веществ из неорганических, хромопласты, содержащие красные, оранжевые и желтые пигменты (каротиноиды), придающие яркую окраску цветкам и плодам растений, лейкопласты, бесцветные пластиды, осуществляющие хранение и мобилизацию запасов питательных веществ. Свободно расположенные клетки обычно имеют округлую форму в виде эллипсоида. Клетки, из которых сложены ткани, контактируют друг с другом по плоским поверхностям и имеют форму многогранника. Число граней может изменяться от 9 до 20. По форме клетки могут быть округлыми, изодиаметрическими, имеющими приблизительно равные диаметры по трем осям. Такие клетки называются паренхимными. В том случае, когда клетки весьма значительно вытянуты, их относят к имеющим прозенхимную форму. Степень удлинения таких клеток характеризуется коэффициентом прозенхимности, который находится через отношение величины их длинной оси к короткой. Клетки, имеющие коэффициент прозенхимности более 3, считаются прозенхимными. В растении клетки могут быть живыми и мертвыми (в стволе древесных растений более 90% клеток мертвые), но они все необходимы для нормальной жизнедеятельности организма. МЕМБРАНЫ КЛЕТКИ Клеточные мембраны (рис. 6) выполняют целый ряд функций: разграничительные, активный перенос различных веществ, пассивное осмотическое перемещение воды, осуществление разнообразных реакций. Клеточные мембраны представляют собой жидкостно-мозаичные системы. Мембраны состоят, в основном, из липидов и белков. Липиды – это органические вещества, которые не растворимы в воде, но вымываются из клеток органическими растворителями – эфиром, хлороформом, бензолом. В большинстве – это сложные эфиры 18 жирных кислот и какого-либо спирта. В клеточных мембранах основу составляет двойной слой фосфолипидов, а также небольшое количество других липидов. Каждая молекула фосфолипида представляет собой глицерол с присоединенными двумя остатками жирных кислот и одним остатком фосфорной кислоты. Глицерол и остаток фосфорной кислоты составляют голову фосфолипида, обладающую полярными, гидрофильными, свойствами. Два неполярных, гидрофобных, хвоста образованы остатками жирных кислот. В мембране липиды повернуты друг к другу своими гидрофобными концами, а гидрофильные головы обращены наружу. Липиды в мембране не остаются жестко закрепленными, а способны перемещаться. Во взаимодействии с липидами в мембране находятся белки, выполняющие структурную роль, а также другие функции. Мембранные белки могут примыкать к бислою липидов, погружаться в него, или пронизывать его насквозь. Белки действуют как активные переносчики различных веществ, образуют гидрофильные каналы, функционируют в качестве катализаторов реакций в виде ферментов, переносят электроны, преобразуют энергию, участвуя в фотосинтезе и дыхании. Гликопротеины – соединения белков и сахаров – участвуют в распознавании внешних сигналов клетки. Мембраны разделяют пространство клетки на большое количество отдельных отсеков – компартментов, где одновременно протекает множество различных реакций метаболизма. Двойные мембранные слои образуют оболочки крупных органелл клетки – ядра, пластид, митохондрий. Одной мембраной окружен весь протопласт клетки, вакуоли, микротельца, каналы эндоплазматического ретикулума, цистерны и пузырьки аппарата гольджи. Рис. 6. Модель биологической мембраны: 1 – туннельный белок; 2 – белковый слой; 3 – двойной липидный слой; 4 – гидрофильная пора Молекулы липидов в двойном липидном слое направлены своими гидрофильными, или полярными, концами (обозначены кружками) во внешние стороны, а гидрофобными неполярными концами внутрь мембраны. Белковые слои состоят из отдельных субъединиц. Некоторые из них (туннельные белки) пересекают всю мембрану. 19 ЦИТОПЛАЗМА Вещество, заполняющее пространство между органеллами, обозначается как основная плазма, гиалоплазма или цитозоль. Она бесструктурная и имеет систему микрофиламентов. Цитозоль содержит около 90% воды, в которой растворены соли, сахара, аминокислоты, жирные кислоты, нуклеотиды и газы. Крупные молекулы – белки и РНК образуют коллоидные растворы. Во внешних слоях протопласта основная плазма более вязкая и по консистенции близка к гелю. В цитозоле не только находятся биомолекулы, но и протекает ряд процессов метаболизма – гликолиз, синтез жирных кислот, нуклеотидов и некоторых аминокислот. Гиалоплазма способна к движению, она перетекает внутри клетки, увлекая при этом органеллы. Это явление называется циклозом. ЯДРО ДНК, ответственная за хранение и передачу информации о структуре белков, локализована в хромосомах. Они заключены в ядро – органеллу, ограниченную мембранами. В ядре содержатся ядрышки – тельца, осуществляющие синтез рибосомальной РНК и формирование предшественников рибосом. Ядра имеются во всех эукариотических клетках, за исключением зрелых члеников ситовидных трубок флоэмы и зрелых эритроцитов млекопитающих. Некоторые специализированные клетки становятся многоядерными (ценоцитными), например двуядерные в ткани, окружающей спорогенные клетки – тапетуме. В одноядерных клетках иногда происходит многократное удвоение ДНК, в результате чего ядро становится полиплоидным, что обозначается как явление эндоплоидии. Ядро играет основную роль в клеточном делении. В период между делениями в интерфазе ядро представляет собой самостоятельную наиболее крупную органеллу клетки, содержащую хроматин и ядрышки, и окруженную оболочкой, состоящей из двух мембран. Внутренний объем ядра заполнен прозрачной коллоидальной средой – кариолимфой, называемой матриксом. В ней погружены ядрышки и хроматин ядра, который в период интерфазы находится в раскрученном деспирализованном состоянии. Более плотная часть хроматина, собранная в виде глыбок, обозначается как гетерохроматин, а рыхлая часть – эухроматин. Хроматин представляет собой нуклеопротеид, включающий ДНК в соединении с гистонами – основными белками. Они образуют нуклеосомы – структуры, напоми20 нающие бусины. В процессе деления клетки хроматин конденсируется в виде хромосом. В ядре также могут находиться белковые включения в виде кристаллов, фибрилл или в бесструктурной аморфной форме. Оболочка ядра сложена двумя мембранами, между которыми имеется перинуклеарное пространство. Наружная мембрана переходит непосредственно в эндоплазматический ретикулум. Ядерная оболочка имеет множество пор. Вокруг каждой поры обе мембраны смыкаются, образуя кольцо (аннулус). В осевой части ядерной поры имеется центральная гранула, окруженная системой фибрилл. Ядро часто содержит по два ядрышка – по одному ядрышку на каждый гаплоидный набор хромосом. В ядрышке происходит синтез рибосомной РНК. В ядрышке можно различить элементы двух типов – гранулярные и фибриллярные, а также вакуоли. Ядрышко связано с определенной частью хроматина – ядрышковым организатором; считается, что это зона вторичной перетяжки хромосомы. В процессе деления ядро претерпевает определенные изменения. Ядро клетки может делиться двумя путями – по типу митоза и мейоза. Во время митоза хромосомы удваиваются, и дочерние клетки получают то же самое количество хромосом, идентичных по составу генов, что и исходная клетка. По типу митоза делятся соматические клетки основной массы тела растения. Во время мейоза дочерние клетки получают половину первоначального числа хромосом, измененных по генетическому составу относительно материнской клетки. Мейоз сопровождается формированием репродуктивных половых клеток. Процессы, происходящие между делением одной клетки и делением клетки следующего поколения, называют клеточным или митотическим циклом. Этот цикл включает ряд последовательных фаз деления ядра: 1) интерфазу, имеющую периоды – пресинтетический G1, синтетический – S и премитотический – G2; 2) профазу; 3) метафазу; 4) анафазу; 5) телофазу. Деление ядра завершается делением клетки – цитокинезом. Помимо копирования всего генетического материала в процессе клеточного деления, ядро контролирует синтез белка в клетке. При снятии копий участков ДНК – транскрипции синтезируется матричная или информационная РНК (м-РНК), которая переносится к рибосомной РНК, находящейся в цитоплазме. Здесь при участии транспортных РНК происходит синтез белков, в основном выполняющих ферментативную функцию. Ферменты ускоряют биологические реакции, а также определяют характер метаболизма и направление развития клетки. 21 ПЛАСТИДЫ Пластиды относятся к типичным органеллам растительных клеток. У высших растений они, как правило, имеют эллипсоидальную форму в виде дисков диаметром 3-10 мкм. В клетке обычно содержится несколько десятков пластид. В зависимости от окраски, связанной с наличием различных пигментов, особенностей функции и структуры, выделяются следующие основные типы пластид: хлоропласты, хромопласты и лейкопласты. Хлоропласты относятся к зеленым пластидам. При участии световой энергии хлоропласты осуществляют процесс фотосинтеза органических веществ из неорганических веществ воды и углекислоты. Эти пластиды содержат зеленые пигменты – хлорофиллы, а также желто-оранжевые каротиноидные пигменты. Хромопласты содержат каротиноиды и присутствуют в окрашенных частях растений – в лепестках цветов, плодах томатов, рябины, корнеплодах моркови др. Лейкопласты представляют собой бесцветные пластиды, осуществляющие запасание веществ. В зависимости от запасаемых веществ лейкопласты подразделяются на амилопласты, накапливающие крахмал, элайопласты – жиры, протеинопласты – белки. При развитии в затененных частях растений также образуются бесцветные пластиды – этиопласты, которые легко способны превращаться в хлоропласты при освещении. Кроме этого, хромопласты и лейкопласты также способны преобразовываться в хлоропласты, например при освещении корнеплодов моркови, клубней картофеля. Пластиды развиваются из небольших бесцветных пропластид, содержащихся в меристематических клетках. В растущей клетке количество хлоропластов увеличивается путем деления. Пластиды имеют сходное строение, отличаясь по развитию внутренней мембранной системы, составу пигментов и накапливаемым веществам. Снаружи пластида окружена оболочкой, состоящей из двух мембран. Внутренний объем заполнен жидкой стромой, в которую погружены плоские мембранные цистерны – тилакоиды. Наиболее сложную тилакоидную систему имеют хлоропласты. У них эта система состоит из стопок округлых коротких цистерн – гран, называемых тилакоидами гран, которые соединяются между собой длинными цистернами – тилакоидами стромы. Между тилакоидами стромы откладываются крахмальные зерна. Пропластиды, лейкопласты и хромопласты не имеют гран. Обычным компонентом стро22 мы пластид являются капли жира – пластоглобулы. Кроме того, в строме содержится кольцевая ДНК и рибосомы, что обеспечивает возможность пластидам самостоятельно синтезировать белки. Это обстоятельство позволило сделать предположение, что в филогенезе пластиды проникли в эукариотические клетки как прокариотические симбионты. МИТОХОНДРИИ Аэробное дыхание осуществляется органеллой – митохондрией. Оболочка митохондрии состоит из двух мембран. Внутренняя мембрана образует складки – кристы. Внутренний объем митохондрии заполнен матриксом, в котором находится одна кольцевая молекула ДНК, небольшое количество рибосом и фосфатные гранулы. При аэробном дыхании в кристах происходит заключительный этап дыхания – окислительное фосфорилирование и перенос электронов. В матриксе функционируют ферменты, участвующие в цикле Кребса и в окислении жирных кислот. Интенсификация дыхания клетки приводит к увеличению объема митохондрий и в них возрастает размер и число крист. Воспроизводятся митохондрии делением, которому предшествует их рост. Существует представление о том, что митохондрии подобно пластидам в филогенезе оказались связанными с эукариотической клеткой в качестве внедрившихся симбионтов. МИКРОТЕЛЬЦА – ПЕРОКСИСОМЫ, ЦИТОСОМЫ Особые органеллы – микротельца – содержат ферменты, необходимые для осуществления различных процессов метаболизма. Микротельца встречаются во многих тканях растений, в особенности в ассимиляционной паренхиме, где взаимодействуют с хлоропластами. Оболочка микротелец образована одной мембраной. Они содержат гранулярный или фибриллярный матрикс, в который погружен один кристалл. Отличительная особенность пероксисом состоит в том, что они содержат фермент каталазу, ускоряющий разложение перекиси водорода на воду и кислород. Пероксисомы листьев расщепляют перекись водорода, возникающую при фотодыхании. Среди цитосом различаются глиоксисомы, участвующие в расщеплении липидов до сахарозы в богатых маслами семенах. 23 ВАКУОЛИ Вакуоли являются важным компонентом протопласта растительных клеток. Они представляют собой полости в цитоплазме, заполненные клеточным соком (водянистой жидкостью). Клеточный сок окружен вакуолярной мембраной – тонопластом, обладающей избирательной проницаемостью. Вакуоли выполняют разнообразные функции в клетке, к которым относятся регуляция водно-солевого обмена, поддержание осмотического потенциала и тургорного гидростатического давления, накопление запасных веществ и консервация отбросов – конечных продуктов метаболизма. В молодых клетках вакуолярная система представлена мелкими вакуолями. В зрелой клетке основной объем занят одной центральной вакуолью. Вакуолярная система клетки формируется из расширенных цистерн эндоплазматического ретикулума, а также путем включения в вакуолярные инвагинации (изгибы) участков цитоплазмы. Вакуолярный клеточный сок содержит водные растворы разнообразных органических и неорганических веществ. К ним относятся запасные соединения: сахара, органические кислоты, белки, фосфатиды. Вакуоли также содержат вещества вторичного обмена: полифенольные соединения – дубильные вещества (таннины), азотсодержащие вещества – алкалоиды, пигменты – антоцианы и конечные продукты жизнедеятельности – соли оксалата или карбоната кальция и др. В вакуолярном соке также содержатся растворенные неорганические соединения. Вакуоли способны разрушать компоненты клетки или запасных веществ. Они участвуют в процессах мобилизации запасных веществ, старения и дифференциации клетки. СТРУКТУРЫ, СВЯЗАННЫЕ С ПЛАЗМАЛЕММОЙ Клеточная мембрана – плазмалемма не всегда плотно примыкает к оболочке клетки, а образует углубления – инвагинации различной формы в протопласте. Эти инвагинации отделяются от плазмалеммы и внедряются в вакуоль в виде множества мультивезикулярных телец. В том случае, когда пузырьки появляются в протопласте в результате деятельности плазмалеммы, их называют плазмалеммасомами, а если они возникают из других мембран клетки – ломасомами. Считается, что наличие инвагинаций плазмалеммы и образование их них пузырьков связано с ростом клеточной оболочки, а также с обменом веществ между протопластом и окружающим его пространством оболочек клеток – апопластом. 24 РИБОСОМЫ Рибосомы представляют собой частицы диаметром 20-30 нм. Они осуществляют синтез белков из аминокислот. Каждая рибосома состоит из двух субъединиц – большой и малой, объединенных ионами магния. Субъединицы образуются в ядрышке, поступают в цитоплазму, где происходит их сборка на молекуле матричной РНК. Рибосомы могут находиться свободно в цитоплазме, прикрепляться к каналам эндоплазматической сети, или объединяться в группы – полирибосомы. В пластидах и митохондриях также содержатся рибосомы, но они отличаются от цитоплазматических рибосом. АППАРАТ ГОЛЬДЖИ Аппарат Гольджи клетки представляет собой совокупность некоторого числа расположенных в цитоплазме телец Гольджи – диктиосом (греч. «диктион» – сеть, «сома» – тело) и выделяемых ими пузырьков Гольджи – везикул. Каждая диктиосома состоит из пачки плоских мембранных цистерн, имеющей регенерационный и секреторный полюсы. На секреторном полюсе цистерны распадаются, начиная от краев, на отдельные пузырьки – везикулы, содержащие полисахариды или полисахаридно-белковые соединения. На регенерационном полюсе из каналов эндоплазматического ретикулума формируются новые цистерны. Везикулы участвуют в построении оболочки клетки, а также в секреции – выделении клеткой веществ. Диктиосомы осуществляют формирование вакуолей клетки. В период цитокинеза диктиосомы концентрируются в экваториальной плоскости клетки и поставляют в виде везикул материал для формирования клеточной перегородки. ЭНДОПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ (ЭР) Эндоплазматический ретикулум представляет собой мембранную систему внутриклеточной циркуляции веществ, а также осуществления ряда биохимических процессов. Эндоплазматическая сеть сложена из системы каналов, пузырьков и цистерн, ограниченных мембраной. На своей внешней поверхности более узкие каналы содержат рибосомы, и эту часть называют шероховатым или гранулярным ретикулумом. Здесь протекают реакции синтеза белков, в том числе и ферментов. Гладкий – агранулярный ретикулум не содержит рибосом и имеет более широкие каналы. В нем образуются углеводы, липиды, терпеноиды. Каналы эндоплазматической сети смежных клеток участвуют в межклеточном обмене веществ, связываясь между собой через плазмодесмы, расположенные в клеточной оболочке. 25 ГЛОБУЛЫ ЛИПИДОВ Это сильно преломляющие свет сферические тельца, называемые сферосомами. Они имеют диаметр до 0,5 мкм и состоят из липидов. Их также называют липидными каплями, которые в электронном микроскопе после фиксации осмиевой кислотой выглядят как темные осмиофильные глобулы. Запасы липидов клетки хранятся в виде сферосом. МИКРОТРУБОЧКИ Микротрубочки расположены в периферическом слое цитоплазмы неделящихся растительных клеток. Наружный диаметр микротрубочек около 24 нм, а внутренний – 14 нм. Микротрубочки расположены параллельно между собой и поперек продольной оси клетки. Они состоят из кислого белка – тубулина, организованного в виде трубки, сложенной из 13 субъединиц. В состав каждой из них входит два глобулярных мономера α и β тубулинов. Микротрубочки принимают активное участие в делении ядра, образуя веретено. Рис. 7. Крахмальные гранулы: 1 – сложные (овес – Avena), 2 – простые и полусложные (картофель – Solanum), 3 – простые (молочай – Euphorbia), 4 – простые (в клетке черешка листа герани – Geranium), 5 – простые (фасоль – Phaseolus), 6 – простые (кукуруза – Zea), 7 – простые крупные и мелкие (пшеница – Triticum) 26 Они прикреплены к первичной перетяжке хромосомы и растягивают хроматиды по полюсам клетки. Микротрубочки осуществляют перемещение пузырьков Гольджи, обеспечивая организованный рост оболочки клетки. При их участии происходит перемещение в клетке больших органелл, например митохондрий, лизосом. МИКРОФИЛАМЕНТЫ Представляют собой очень тонкие нити диаметром 5-7 нм, состоящие из белка актина, аналогичного тому, который имеется в мышцах. Микрофиламенты образуют наряду с микротрубочками цитоскелет клетки. Они осуществляют передвижение цитоплазмы и пространственную организацию обмена веществ в цитоплазме. ЭРГАСТИЧЕСКИЕ ВЕЩЕСТВА КРАХМАЛ. Первичный крахмал возникает в хлоропластах из продуктов фотосинтеза. Вторичный, запасной, крахмал накапливается в лейкопластах. Каждая крахмальная гранула имеет образовательный центр и расположенные вокруг него концентрические слои роста (рис. 7). Слои роста образуются за счет чередования углеводов амилозы с линейными молекулами, мономером которых является α-d-глюкопираноза, и амилопектина с разветвленными молекулами, состоящими из фрагментов амилозы, менее растворимым в воде. Углеводы могут запасаться в клетках также в виде инулина – полимера d-фруктозы. Он откладывается в клетках в форме сферокристаллов, образованных из множества игловидных кристаллов (рис. 8). При своем росте эти сферокристаллы могут захватывать несколько клеток. БЕЛКИ. Простые белки – протеины накапливаются в клетке в виде алейроновых зерен – твердых телец (рис. 9). Чаще всего белки накапливаются в семенах – в наружном алейроновом слое эндосперма зерновок злаковых, в семядолях бобовых. Запасные Рис. 8. Запасающая паренхима в клубне топинамбура (Helianthus tuberosus): 1 – сферокристаллы инулина 27 белки накапливаются в вакуолях и по мере потери влаги выпадают в осадок в виде зерен округлой или эллипсоидальной формы. Они окружены элементарной мембраной – тонопластом. Простые алейроновые зерна не имеют внутренней структуры. В сложных алейроновых зернах аморфный матрикс окружает одно или несколько кристаллоподобных включений – кристаллоидов или аморфные тельца округлой формы – глобоиды. Рис. 9. Запасной белок в клетках эндосперма семени клещевины (Ricinus communis): 1 – аморфная белковая масса – матрикс, 2 – глобоид, 3 – алейроновое зерно, 4 – оболочка алейронового зерна, 5 – кристаллоид ДУБИЛЬНЫЕ ВЕЩЕСТВА – ТАННИДЫ. Таннины относятся к высокомолекулярным фенольным соединениям. Они обладают способностью осаждать белки и алкалоиды и имеют вяжущий вкус. Танниды встречаются в вакуолях, гиалоплазме в виде телец различных размеров желтого, красного или коричневого цвета, или тонкогранулярных скоплений. Эти вещества присутствуют в листьях, проводящих тканях, перидерме, семенной кожуре, незрелых плодах, патологических образованиях – галлах. Танниды также накапливаются в коре и ядровой части древесины некоторых пород. ЖИРЫ, МАСЛА И ВОСКИ. Воски – это сложные смеси, состоящие из эфиров жирных кислот и одноатомных или двухатомных спиртов. Масла и жиры относятся к глицеридам органических кислот и принадлежат к запасным формам липидов. Масла представляют собой жидкие вещества, а жиры – твердые. Липиды чаще всего встречаются в клетке в форме капель. Жиры синтезируются в цитоплазме, а также в пластидах – элайопластах. Эфирные масла имеют низкий молекулярный вес. Они накапливаются в специализированных клетках – идиобластах, либо в межклеточных полостях – вместилищах. ТЕРПЕНОИДЫ. Это обширный класс природных органических соединений, относящихся к вторичным метаболитам, с общей формулой (С5Н8)n, где n ≥ 2. Их также обозначают как изопреноиды, или 28 соединения изопентиловой группы, поскольку они построены из остатков непредельного углеводорода изопрена: CH2 = С(СН3) – CH = CH2. Терпеноиды обнаружены практически во всех тканях растений и содержатся в эфирных маслах, скипидаре, смолах, бальзамах. Производными изопрена являются каротиноиды, каучук, гуттаперча. Секреция терпеноидов осуществляется одноклеточными образованиями – масляными клетками, секреторными идиобластами, нечленистыми млечниками или многоклеточными структурами – железистыми волосками, железками, эпителием смоляных ходов и вместилищ. Синтез терпеноидов осуществляется в агранулярном эндоплазматическом ретикулуме терпеноидогенных клеток. КРИСТАЛЛЫ. В вакуолях растительных клеток в значительных количествах накапливаются кристаллы оксалата или карбоната кальция. В большинстве случаев кристаллические скопления этих соединений образуются как конечные продукты жизнедеятельности протопласта для выведения их из обмена веществ. Поэтому данные кристаллы концентрируются в тканях и органах, которые периодически сбрасываются растениями, например, в листьях и коре. Форма кристаллов разнообразная (рис. 10). Они могут быть одиночными призматическими, в виде рафид – пучков игольчатых кристаллов, друз – шаровидных скоплений призматических кристаллов, стилоидов – удлиненных колонковидных кристаллов, кристаллического песка – скоплений мелких кристаллов. Рис. 10. Различные типы кристаллов оксалата кальция (щавелевокислого кальция): 1 – призматические одиночные и объединенные крестообразно в клетках сухой чешуи луковицы лука (Allium cepa), 2 – последовательные стадии формирования друз кристаллов в клетках черешка листа бегонии (Begonia manicata), 3 – рафида в клетках корневища купены (Polygonatum officinale) 29 ОБОЛОЧКА КЛЕТКИ Растительная клетка характеризуется жесткой клеточной оболочкой, имеющей различный состав и свойства в зависимости от местоположения клетки в растении и выполняемых ею функций. Одно из основных назначений клеточной оболочки – обеспечение поддержания устойчивой формы и механической прочности клеткам и органам растения. Опорная функция усиливается благодаря дополнительному увеличению толщины оболочки. Прочность тонких оболочек на растяжение обеспечивает поддержание устойчивой формы молодых, а также слабо одревесневших органов за счет упругости клеток – их тургорного состояния. Оно создается внутренним гидростатическим давлением протопласта на клеточную оболочку. Оболочка молодых, а в определенных условиях и зрелых клеток (например, при ее подкислении), обладает некоторой пластичностью. Это свойство обусловливает возможность роста оболочки растяжением под действием тургора. Рост клеток приводит к росту органов, в состав которых они входят. Направление роста клеток и их форма предопределяются характером отложения в клеточной оболочке микрофибрилл целлюлозы. Если микрофибриллы ориентированы параллельно между собой и поперек продольной оси органа, то рост таких клеток происходит в направлении данной оси. Клетки при этом приобретают вытянутую форму, не изменяясь существенно по размерам в поперечном направлении. Оболочка растительной клетки проницаема для воды и низкомолекулярных веществ. Это свойство обеспечивает возможность транспорта веществ по апопласту – системе оболочек взаимосвязанных между собой клеток. В семенах оболочка клеток выполняет функцию запасания питательных веществ. Оболочка, пропитанная жироподобными веществами, препятствует передвижению по ней воды и растворенных веществ. Водонепроницаемая оболочка формируется у клеток покровных тканей, а также в тканях, выполняющих барьерную функцию, у которых избирательный транспорт веществ направляется только через протопласты клеток, например в эндодерме. Древесина состоит, главным образом, из оболочек клеток. Она занимает наибольший объем в стволах и нередко образует основную продукцию древесных растений. Поэтому сведения о структуре, составе и свойствах оболочки растительных клеток имеют большое 30 значение для специалистов, занимающихся лесоводством, заготовкой, переработкой древесины, изготовлением и эксплуатацией древесных конструкций, их защитой от разрушения. Оболочка клетки представляет собой продукт жизнедеятельности ее протопласта. Она функционирует как при жизни клетки, так и после ее отмирания, сохраняя механическую, проводящую и защитную функции. Структура и состав клеточной оболочки Оболочка зрелой растительной клетки имеет слоистое строение. Центральный слой между двумя смежными клетками составлен срединной пластинкой, состоящей из пектиновых веществ (рис. 11). К ней примыкает первичная оболочка. Обычно она не имеет большой толщины и обладает значительной пластичностью, не препятствуя росту клетки. У клеток некоторых тканей, например колленхимы, первичная оболочка значительно утолщается в отдельных частях клеточной стенки. Это придает им дополнительную механическую прочность, при сохранении определенной пластичности. К внутренней стороне первичной оболочки, обращенной к полости клетки, примыкает вторичная оболочка, которая часто приобретает значительную толщину. Вторичная оболочка подразделяется на три слоя – S1, S2 и S3, счет которых начинается от первичной оболочки. Слой S2 – самый толстый из них. У волокон и трахеид древесных растений внутренняя поверхность слоя S3, покрыта бородавчатым слоем, состоящим из нецеллюлозной пленки с бугорками, представРис. 11. Поперечный срез ляющим собой остатки протооболочек растительных клеток, пласта. Этот слой иногда относят расположенных в ткани: к третичной оболочке. 0 – срединная пластинка; Оболочка клетки имеет констI – первичная оболочка; рукцию в виде каркаса, сложенII – вторичная оболочка, состоящая ного из мицелл целлюлозы, произ трех слоев – S1, S2 и S3; странство между которыми заIII – третичная оболочка, к которой полнено матриксом (рис. 12). относится бородавчатый слой 31 Рис. 12. Ультраструктура клеточной оболочки. I – оболочка без включений, состоящая из матрикса и целлюлозного каркаса; II – оболочка, инкрустированная лигнином; III – оболочка, инкрустированная лигнином, а также полифенолами или минеральными веществами. 1 – матрикс, 2 – целлюлозный каркас, 3 – лигнин, 4 – полифенолы, минеральные вещества Поверхность целлюлозного каркаса инкрустируется лигнином при одревеснении оболочки либо суберином при ее опробковении. Целлюлоза является основным скелетным веществом клеточных оболочек практически всех растений. Ее состав описывается формулой (С6Н10О5)n (рис. 13). Целлюлоза образуется путем полимеризации в линейную неразветвленную цепь некоторого количества молекул β-глюкозы (от нескольких сотен до нескольких тысяч). Целлюлоза представляет собой очень стойкое вещество, ее гидролиз осуществляется только сильными кислотами. Целлюлозный каркас оболочки клетки состоит из системы фибрилл (см. рис. 13). Макрофибриллы целлюлозы видны в световом микроскопе. Микрофибриллы можно наблюдать в электронном микроскопе, их диаметр колеблется у разных клеток в пределах 10–30 нм, а длина может достигать нескольких микрометров. Субъединицы микрофибрилл – мицеллы еще мельче и имеют упорядоченное расположение молекул целлюлозы. Нитевидные молекулы целлюлозы в оболочке клетки располагаются параллельно друг другу и группируются по нескольку десятков, образуя вытянутую трехмерную решетку, характерную для кристаллов. Двойное лучепреломление света оболочками клеток, видимое в поляризационном микроскопе, связано с кристаллическим состоянием молекул целлюлозы в микрофибриллах. Микрофибриллы эластичны и обладают очень высокой механической прочностью на разрыв (соизмеримой со сталью). Микрофибриллы целлюлозы во вторичной оболочке имеют различную ориентацию (рис. 14). В слоях S1 и S3 32 Рис. 13. Детальная структура оболочки растительных клеток. А. Тяж волокнистых клеток. Б. Поперечный срез волокон. В. Участок среднего слоя вторичной оболочки с макрофибриллами целлюлозы (белые включения) и межфибриллярным пространством, заполненным нецеллюлозными веществами (черные включения). Г. Участок макрофибриллы, состоящий из микрофобрилл (белые включения) и пространством между ними, заполненным нецеллюлозными веществами (черные включения). Д. Микрофибриллы, состоящие из цепочек молекул целлюлозы. Участки упорядоченно расположенных молекул целлюлозы образуют мицеллы. Е. Часть мицеллы, в которой цепочки молекул целлюлозы образуют пространственную решетку. Ж. Два остатка глюкозы, связанные между собой атомом кислорода представляют собой часть молекулы целлюлозы. 1 – срединная пластинка; 2 – первичная оболочка; 3 – трехслойная вторичная оболочка; 4 – макрофибрилла; 5 – микрофибрилла; 6 – молекула целлюлозы; 7 – мицеллы микрофибриллы расположены в виде спиралей, витки которых образуют большой угол с продольной осью клетки, что обеспечивает эластичность этих слоев по данной оси. В слое S2 микрофибриллы образуют витки спирали, которые располагаются под малым углом к 33 продольной оси клетки, препятствуя ее растяжению в этом направлении. Целлюлоза взаимосвязана с матриксом оболочки, в состав которого входят нецеллюлозные полисахариды – гемицеллюлозы и пектиновые вещества – полиурониды. Рис. 14. Схематическое изображение слоев оболочки трахеиды и направление отложения в них микрофибрилл: 1 – бородавчатый слой; слои вторичной оболочки; 2 – S3, 3 – S2, 4 – S1; 5 – первичная оболочка; 6 – срединная пластинка К гемицеллюлозам относятся нецеллюлозные гексозаны – маннан и галактан, характеризующиеся формулой (С6Н10О5)n, а также пентозаны – ксилан и арабан, описываемые общей формулой (С5Н8О4)n. У маннана молекула состоит из 7080 единиц маннозы. Молекула галактана состоит из 8-10 единиц галактозы. Молекулы ксилана и его изомера арабана состоят приблизительно из 20 единиц ксилозы или арабинозы. Кроме этого в клеточных оболочках присутствуют гексозанпентозаны или гексопентозаны, такие как глюкоарабаны, глюкоксиланы, галактоарабаны и галактоксиланы, строение которых еще полностью не установлено. Гемицеллюлозы разделяют на две основные группы. 1. Гемицеллюлозы, прочно связанные с целлюлозой; они не отделимы от нее простыми методами. К ним относятся пентозаны, ксилан и маннан. 2. Гемицеллюлозы, связанные с целлюлозой свободными связями, так называемые свободные гемицеллюлозы. Они делятся на простые, к которым относятся арабан, ксилан, маннан, галактан и гексопентозаны, а также на гемицеллюлозы на основе уроновых кислот, чаще всего глюкуроновой. Уроновые кислоты образуют цепочки молекул различной длины и в соединении с гексозанами или пентозанами гемицеллюлоз дают полиурониды. Пектиновые вещества образуют центральный клеящий слой – срединную пластинку, общий для двух соседних оболочек клеток. К пектиновым веществам относится группа веществ, близких по составу: 1) пектиновая кислота; 2) пектаты (соли пектиновой кислоты), пектаты Na и K, растворимые в воде, а также пектаты Ca и Mg, не растворимые в воде; 3) сложные эфиры пектиновой кислоты, рас34 творимые в воде, называемые пектином, и не растворимые в воде, называемые пектозой. Пектины состоят из галактуроновой кислоты, образующей длинные цепи полигалактуроновых кислот, часто метилированных. Им сопутствуют постоянные количества l-арабинозы и d-галактозы. Часть молекулы полигалактуроновой кислоты В состав оболочки клетки также входят неуглеводные компоненты: лигнин, являющийся полимером полифенольной природы, а также липиды, к которым относятся воски, кутин и суберин. Лигнин представляет собой комплекс веществ, пропитывающих целлюлозные клеточные оболочки и вызывающих их одревеснение. Существует представление о том, что возникновение лигнина у растений способствовало их переходу от водного существования к наземному. Химическое строение лигнина и его свойства различны на разных стадиях одревеснения и в зависимости от вида древесных пород. Лигнин, выделенный любым способом, отличается по строению и свойствам от того, который находится в целлюлозной клеточной оболочке. Исходными веществами в образовании лигнина покрытосеменных растений являются простые глюциды – гексозы и пентозы с присоединенными метоксильными группами (-О.СН3), а также пектиновые вещества. У хвойных растений в процессе одревеснения метилирование гексоз и пентоз приводит к синтезу глюкозида – кониферина. Затем фермент глюкозидаза разлагает кониферин и синтез лигнина начинается из освобожденного кониферилового спирта: Из этого вещества также синтезируются таннины и смолы. Лигнин представляет собой аморфное вещество. После освобождения от целлюлозы клеточной оболочки лигнин приобретает коричневую или черно-бурую окраску. Например, в результате разложения целлюлозы целлюлозуразрушающими грибами и освобожде35 ния лигнина при бурой гнили древесина приобретает коричневую окраску. Опробковение вторичной оболочки клеток происходит в результате отложения в ней суберина. Такая оболочка становится непроницаемой для воды и газообразных веществ. Суберин представляет собой высокополимерное липидное соединение, состоящее из жирных кислот. На внешней поверхности органов растения, соприкасающихся с воздухом, для их защиты от излишнего испарения воды также откладываются липидные вещества кутины и воска. Оболочки клеток могут пропитываться кремнеземом, что придает им дополнительную механическую прочность. Спорополленинами защищается оболочка пыльцевых зерен и спор грибов, что придает им исключительную стойкость к факторам внешней среды, поскольку эти вещества наиболее устойчивые из всех известных органических соединений. Благодаря такому свойству спорополленина, пыльцевые зерна и споры грибов сохраняются в торфяных залежах в течение сотен веков. У водных растений на внешней поверхности оболочек эпидермальных клеток, обращенных в водную среду, откладываются гидрофильные слизистые вещества. Ослизнение оболочек полисахаридными слизями происходит у клеток корневого чехлика, а также эпидермы семян некоторых растений, например льна при увлажнении. На внутренней поверхности оболочек различных типов клеток, в том числе ситовидных трубок флоэмы, пыльцевых трубок, а также гиф грибов, происходит отложение полисахарида каллозы. Это вещество осуществляет закупоривание ситовидных трубок к моменту прекращения их активной деятельности. В состав оболочки растительной клетки входят также белки, вода и минеральные вещества. Формирование клеточной оболочки Клеточная оболочка начинает формироваться в конце процесса деления ядра клетки (кариокинеза). После расхождения хромосом в анафазе в делящейся клетке формируется фрагмопласт (от греч. «фрагма» – изгородь, разделение). Первоначально он представляет собой густую цитоплазматическую массу с микротрубочками в виде веретена (рис. 15). Затем фрагмопласт уплощается и приобретает форму двояковыпуклой линзы. При этом оба дочерних ядра делящейся клетки смещаются к ее середине. Первым признаком возник36 новения клеточной оболочки служит появление в экваториальной плоскости между микротрубочками фрагмопласта мелких пузырьков аппарата Гольджи. Они сливаются, образуя клеточную пластинку, которая представляет собой первичную структуру клеточной оболочки. Это свидетельствует о начале процесса цитокинеза – разделения материнской клетки на две дочерние. Клеточная пластинка разрастается по направлению к периферии материнской клетки, достигает ее продольной стенки и сливается с нею. Рост клеточной пластинки происходит путем включения в нее дополнительных пузырьков аппарата Гольджи (рис. 16). Рис. 15. Образование клеточной пластинки во время митоза. I – ранняя телофаза. В экваториальной плоскости фрагмопласта между микротрубочками начинается формирование клеточной пластинки из пузырьков Гольджи. II – поздняя телофаза. По мере центробежного роста клеточной пластинки, фрагмопласт укорачивается и расширяется в экваториальной плоскости, достигая противоположных стенок материнской клетки. 1 – хромосомы одной из дочерних клеток, 2 – границы фрагмопласта, 3 – микротрубочки, 4 – клеточная пластинка, формируемая пузырьками Гольджи, 5 – два ядра дочерних клеток, 6 – оболочка материнской клетки Пузырьки содержат кислые компоненты, такие как пектиновые вещества, а также другие соединения уроновых кислот, играющие роль матрикса в процессе синтеза будущей клеточной оболочки. Элементарная мембрана, окружающая пузырьки, при их слиянии включается в плазмалемму двух разделяющихся протопластов дочерних клеток. В процессе роста клеточной пластинки формируются плазмодесмы. Каждая плазмодесма представляют собой канал в клеточной 37 пластинке, выстланный плазмалеммой. По оси канала расположена десмотрубочка, стенки которой являются продолжением каналов эндоплазматического ретикулума смежных клеток. Таким образом, у дочерних клеток по плазмодесмам оказываются связанными между собой гиалоплазма и каналы эндоплазматического ретикулума. Система плазмодесм образует между живыми клетками единое целое – симпласт. По симпласту между клетками транспортируются ионы, молекулы и передается возбуждение. Рис. 16. Формирование клеточной пластинки в период цитокинеза. I – слияние пузырьков аппарата Гольджи и образование клеточной пластинки и плазмодесм; II – продольный разрез через плазмодесму с ответвлением канала эндоплазматической сети; III – поперечный разрез через плазмодесму; IV – широкая пора с несколькими каналами эндоплазматической сети в ситовидной трубке флоэмы. 1 – тельце Гольджи (диктиосома); 2 – пузырек Гольджи; 3 – слияние пузырьков Гольджи; 4 – примыкание участков клеточной пластинки; 5 – новая плазмалемма, образующаяся из мембран пузырьков Гольджи; 6 – клеточная пластинка; 7 – канал эндоплазматической сети, проходящий через плазмодесму; 8 – канал эндоплазматической сети одной из дочерних клеток; 9 – слой цитозоля между плазмалеммой и каналом эндоплазматической сети в плазмодесме; 10 – замыкающая пленка поры Растущая клеточная пластинка, еще до ее соприкосновения с продольной клеточной оболочкой материнской клетки, приобретает трехслойное строение. Центральный слой представляет собой срединную пластинку. Она изотропная и не обладает двойным лучепреломлением. Дочерние клетки, каждая в свою очередь, начинают формировать около срединной пластинки анизотропную первичную 38 оболочку, обладающую двойным лучепреломлением. Это связано с тем, что первичная оболочка, в отличие от срединной пластинки, строится из регулярно расположенных структурных элементов. Клеточная пластинка разрастается, достигает продольных стенок материнской клетки и сливается с ними. Она разделяет полностью между собой две дочерние клетки и превращается в новую клеточную оболочку. Эта оболочка состоит из срединной пластинки, к которой с обеих сторон, обращенных к двум молодым клеткам, примыкают два слоя первичной оболочки. Эти слои первичной оболочки представляют собой продукт жизнедеятельности молодых дочерних клеток. Каждая клетка формирует около срединной пластинки свой слой первичной оболочки. Процесс роста первичной клеточной оболочки продолжается до тех пор, пока клетка сохраняет способность увеличивать свой объем. Наряду с этим в данный период на поверхности первичной оболочки протопластом начинается формирование отдельных участков вторичной оболочки. В молодых растущих органах клетки сосудов и трахеид первоначально формируют редко расположенные одревесневшие кольчатые вторичные утолщения оболочки. Затем формируются близко расположенные кольца. Далее утолщения приобретают вид спиралей. В период завершения роста дифференцирующихся клеток вторичная оболочка начинает приобретать вид жесткой конструкции. Смыкание утолщенных спиралей приводит к образованию лестничных утолщений, которые далее сменяются сетчатыми и пористыми (рис. 56). Рост клеточной оболочки осуществляется путем аппозиции – наложения нового материала на прежние слои. Кроме того, рост оболочки происходит в результате интусуссцепции – включения нового материала, например лигнина или кутина, в уже имеющиеся структуры. Утолщение вторичной оболочки приводит к уменьшению объема полости клетки. Поры клеточной оболочки Взаимосвязь между клетками осуществляется через поры – углубления во вторичной оболочке (рис. 17). Эти углубления обычно расположены друг против друга во вторичных оболочках соседних клеток. Такие углубления образуют две поровые камеры или каналы, разделенные тонкой частью оболочки, называемой замыкающей пленкой поры. 39 В замыкающей пленке поры или поровой мембране имеются плазмодесмы, через которые обеспечивается взаимосвязь цитоплазмы соседних клеток. Каждая плазмодесма представляет собой канал, выстланный плазмалеммой, в центре которого расположена десмотрубочка. Через нее цистерны ЭР соседних клеток сообщаются между собой. При отмирании клеток плазмодесмы исчезают. Пора, имеющая две цилиндрические камеры во вторичной оболочке, называется простой парой пор (рис. 18). Пора называется слепой тогда, когда она находится против межклеточного пространства и не дополняется структурами соседней клетки. Существует окаймленные поры, у которых камеры образованы нависающей над замыкающей пленкой поры вторичной оболочкой (рис. 19). При этом наружный диаметр отверстия камеры значительно меньше внутреннего. У голосеменных растений в замыкающей пленке окаймленных пор трахеид имеется утолщение средней части, называемое торусом. Краевая маргинальная часть замыкающей пленки поры – тонкая и эластичная. Поэтому при возникновении определенной разности давления между соседними клетками Рис. 17. Строение различных типов пор в оболочках клеток. А. Простые поры. Б. Окнообразная пора в клетках радиальных лучей видов рода Pinus. В. Окаймленная пора трахеид и сосудов. Г. Зубчатая пора в оболочке клеток ксилемы некоторых широколиственных древесных пород. 1 – поровая мембрана с плазмодесмами; 2 – камера поры; 3 – срединная пластинка; 4 – вторичная оболочка; 5 – первичная оболочка; 6 – краевая, или маргинальная зона; 7 – торус; 8 – наружное отверстие поры – апертура; 9 – поровая камера 40 Рис. 18. Простые поры в клетках с утолщенной вторичной оболочкой. I – каменистые клетки плода айвы; II – лубяные волокна табака. 1 – простые пары пор, 2 – ветвистые поры, 3 – слепая пора, 4 – межклетник Рис. 19. Схемы строения окаймленных и полуокаймленных пар пор. А. Две окаймленные поры на виде сбоку. Б. Вид с поверхности на окаймленную пору. В. Окаймленная пара пор в закрытом состоянии. Г. Вид с поверхности на полуокаймленные поры. Д. Вид сбоку на полуокаймленные поры. 1 – торус, 2 – маргинальная зона замыкающей пленки поры, 3 – окаймление вторичной оболочки, 4 – камера поры, 5 – наружная апертура поры, 6 – внутренняя апертура поры торус смещается к одному из окаймлений и закрывает узкое отверстие поры. Такая пора оказывается закрытой и не способна пропускать водные растворы. Это свойство окаймленных пор обеспечивает предохранение растений от потери ксилемного со41 ка в случае механического повреждения тканей. Окаймленные поры могут препятствовать различного рода химической обработке древесины хвойных, например их пропитке антисептирующими или другими растворами, либо красителями. Поры могут быть также полуокаймленными в том случае, когда одна камера – окаймленная, а вторая – простая (рис. 19). Кроме того, существуют окаймленные поры со щелевидными камерами, расположенными под углом друг к другу. Межклетники В органах растения между клетками возникают свободные пространства – межклетники. Они могут оставаться в виде системы газообмена между клетками, либо заполняться продуктами жизнедеятельности клеток, образуя системы вместилищ. По способу возникновения межклетники подразделяются на схизогенные и лизигенные. Схизогенные межклетники возникают путем разъединения клеток по срединной пластинке оболочки. Лизигенные межклетники образуются путем растворения лизиса некоторой части клеток. Иногда схизогенный путь формирования межклетников сочетается с лизигенным. Мацерацией называется искусственное разделение клеток какихлибо тканей путем разрушения их срединной пластинки при помощи кислот, ферментов и других веществ. Мацерация применяется при изготовлении анатомических и гистологических препаратов, а также при производстве бумаги, выделении технических волокон из стеблей льна, конопли и т.д. 42 Г л а в а 3. Т К А Н И Ткани представляют собой системы клеток, сходные по строению, функциям и происхождению. Все высшие растения состоят из тканей. Ткань относится к простой в том случае, когда она образована морфологически и функционально одним типом клеток. Существуют также сложные комплексные ткани, состоящие из нескольких типов клеток. В зависимости от выполняемой функции ткани подразделяются на несколько групп: образовательные (меристемы), покровные, проводящие, механические, основные, выделительные (секреторные). Способность к делению – характерная черта клеток меристематических тканей. Все остальные ткани относятся к постоянным, они развиваются из меристем. В процессе дифференциации меристематические клетки, имеющие одинаковое происхождение, приобретают индивидуальные функциональные и морфологические отличия, характерные для постоянных тканей. Это сложный процесс, связанный с действием различных факторов, в том числе и ростовых веществ – гормонов. Тканевая специализация клеток представляет актуальную проблему современной биологии. Дифференцированные клетки, имеющие живые протопласты с полным набором органелл, в определенных условиях вторично приобретают способность к делению. ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ Уникальное свойство неограниченного роста растений обусловлено постоянным присутствием у них молодых тканевых зон – меристем, клетки которых сохраняют способность к делению длительное время. Обычно клетки меристем делятся по типу митоза. В результате такого типа деления дочерние клетки наследуют от материнской клетки идентичные геном и набор хромосом. В зависимости от положения в растении различают несколько типов меристем: 1) апикальные, расположенные на верхушках побегов и окончаниях корней; 2) латеральные, занимающие боковое по43 ложение в органах; 3) вставочные или интеркалярные, которые находятся в основаниях листьев и междоузлий стеблей (например, у злаков); 4) раневые или каллусные, возникающие в местах поранения органов растения. По происхождению в растении выделяют первичные и вторичные меристемы. Апикальные меристемы первичны, они осуществляют формирование новых органов растения и обеспечивают его рост главным образом в длину. Латеральные меристемы обусловливают рост органов по толщине в радиальном направлении. К первичным латеральным меристемам относятся прокамбий и перицикл, а к вторичным – камбий и феллоген. Прокамбий формирует первичные проводящие ткани флоэму и ксилему, а также дает начало развитию камбия. Перицикл расположен обычно в корне под первичной корой, в нем начинается формирование апексов боковых корней; из перицикла формируется феллоген и отчасти камбий. Камбий формирует вторичные проводящие ткани – ксилему и флоэму. Феллоген образует вторичную покровную ткань – перидерму. Интеркалярные меристемы первичны и формируют комплекс тканей стеблей и листьев, в основании которых они располагаются. Раневые меристемы вторичны, они осуществляют формирование однородных паренхимных клеток, защищающих места повреждения тканей и органов растения. Каллусообразование используется в практике вегетативного размножения растений прививками и черенками. АПИКАЛЬНАЯ МЕРИСТЕМА ПОБЕГА Зачаток нового побега формируется клетками апикальной меристемы, расположенной в почке зародыша или материнского побега предыдущей генерации. Как правило, апикальная меристема организована в виде простого бугорка полусферической или конической формы, называемого апексом. Меристематические клетки имеют различия по функциям и морфологии в зависимости от положения в апексе. В связи с этим в апексе побега удается выделить несколько гистологических зон, состоящих из однотипных клеток. 44 Инициаль, или инициальная клетка – это клетка, которая делится на две сестринские клетки, одна из которых остается в меристеме, а другая добавляется к меристематическим тканям, дифференцирующимся, в конечном итоге, в разнообразные ткани, характерные для растения. Клетка, остающаяся в апикальной меристеме, функционирует подобно своей предшественнице как инициаль. Число инициалей в апексах корня и побега варьирует. У споровых (криптогамных) растений в апексе имеется одна или несколько инициальных клеток (рис. 20). В целом апекс у этих растений имеет относительно простое строение. Рис. 20. Апикальные меристемы побегов у несеменных растений. Апикальные инициальные клетки пирамидальной (А) и линзообразной (Б) форм. Дочерние клетки отделяются по трем граням у пирамидальной инициали и по двум – у линзообразной. Апикальные инициальные клетки на продольных срезах побега хвоща (Equisetum) (В) и корневища папоротника орляка (Pteridium) (Г). На рисунке в видны также апикальные клетки листовых примордиев; одна из них (левая) делится. 1 – производные клетки апикальных инициалей; 2 – апикальные инициальные клетки; 3 – листовой примордий Апексы побегов голосеменных растений Достаточно сложную гистологическую структуру имеют апексы побегов голосеменных растений. Впервые наиболее подробно структура апексов побегов голосеменных изучена в 1938–1941 годах А. Фостером. Разработанная им система гистологической зональности апексов голосеменных обозначается «типом гинкго» и широко используется многими исследователями. 45 Рис. 21. Схема строения апекса и зачатка побега в вегетативной почке ели, построенная на основе изучения апикальных меристем ели сибирской – Picea obovata, ели Шренка – Picea schrenkiana и ели обыкновенной – Picea abies. 1 – апикальные инициали, 2 – центральные материнские клетки, 3 – протодерма, 4 – периферическая меристема, 5 – прокамбиальные клетки, 6 – сердцевинная меристема; 7 – листовые зачатки, 8 – прокамбиальные пучки, 9 – диафрагма, 10 – кроющие чешуи почки Так, например, у ели сибирской, ели Шренка, ели обыкновенной выявлена следующая гистологическая зональность апекса побега (рис. 21). На самой верхушке в терминальной части апекса расположена зона апикальных инициальных клеток. Они сложены в один слой. Эти клетки в результате своего деления осуществляют формирование, инициирование нижележащих зон апекса. Главное свойство данных клеток состоит в том, что они остаются продолжительное время (в течение многих лет) в молодом, ювенильном, состоянии и сохраняют свою способность к делению. В апикальных инициалях в наибольшей мере по числу и объему накапливаются сферосомы. Из апикальных инициалей формируется внешний слой клеток апекса – протодерма. Она окружает все возникающие из апекса боковые бугорки – зачатки кроющих чешуй почки и листьев будущего охвоенного побега. В последующем при развитии этих органов из протодермы формируется первичная покровная ткань – эпидерма. Под апикальными инициалями расположена зона центральных материнских клеток. Эти клетки находятся в области слоев 2–6. Они самые крупные в верхушке апекса, диаметром 21–23 мкм, с массивными ядрами в поперечнике 16–17 мкм. Центральные материнские клетки имеют наибольший парциальный объем ядер и самую значительную деспирализацию хроматина по сравнению с остальными клетками апекса. Это обусловливает их высокий метаболизм и возможность активного участия данной зоны в регуляции процессов 46 гистогенеза апекса. Пластиды центральных материнских клеток отличаются низким парциальным объемом, признаками ювенильности структуры. Оболочки данных клеток утолщенные. Центральные материнские клетки осуществляют реализацию генетической программы на морфологическом уровне, они играют роль организатора органогенной деятельности апекса. Центральные материнские клетки в направлении по оси апекса формируют сердцевинную меристему. Она представляет собой зону, которая предопределяет, детерминирует число осевых рядов клеток паренхимы сердцевины будущего стебля побега. Сердцевинная меристема служит местом резервирования запасных веществ в апексе. Это обусловлено активным развитием пластид, которые выполняют в ней роль амилопластов, накапливающих крахмал, а также максимальной вакуолизацией клеток. Клетки сердцевинной меристемы уплощены поперек оси апекса, к его основанию они укрупняются. Эти клетки имеют низкую величину парциального объема ядер и наиболее конденсированное состояние хроматина, что свидетельствует об их относительно небольшой метаболической активности. Часть клеток сердцевинной меристемы приобретает увеличенный объем и накапливает таннины. В процессе развития зачатка побега и продольного роста стебля каждая клетка сердцевинной меристемы формирует колонку, состоящую из нескольких (до 8–10) дочерних клеток, расположенных вдоль оси стебля. Колонка формируется путем деления исходной материнской клетки сердцевины перегородками поперечными оси стебля. В результате такого организованного деления клеток многочисленных колонок сердцевинной меристемы возникают условия осевого роста стебля без существенного увеличения его диаметра. Некоторые авторы называют сердцевинную меристему также колончатой меристемой. Вокруг сердцевинной меристемы центральные материнские клетки вычленяют слой клеток прокамбия. Деление его клеток в основании апекса приводит к образованию прокамбиальных пучков в листовых бугорках и проводящем цилиндре, расположенном на внешней границе сердцевинной меристемы зачатка побега. Клетки прокамбия также, вероятно, осуществляют доставку веществ в апексе. Это обусловлено тем, что прокамбиальные клетки непосредственно связаны с сетью прокамбиальных пучков центрального цилиндра зачатка побега, расположенного ниже апекса. Эта сеть в свою очередь контактирует с камбием проводящего цилиндра укороченного стебля почки. Камбий почки окружен нормально развитыми проводящими элементами флоэмы и ксилемы, которые связаны со стеблем материнского побега. Таким образом, прокамбиальные пучки имеют баланс метаболитов аналогичный тому, который имеет камбий. Транспортная функция прокам47 биальных клеток также обусловлена их удлиненной формой в периклинальном направлении апекса и значительной степенью вакуолизации. Прокамбий наряду с транспортировкой веществ играет роль первичной боковой, латеральной меристемы. В процессе роста структур зачатка побега прокамбий формирует проводящие ткани – первичную флоэму и первичную ксилему. Из прокамбия также формируется вторичная боковая образовательная ткань – камбий. Основную органогенную функцию в апексе осуществляет периферическая меристема, расположенная между протодермой и клетками прокамбия. В терминальной, верхней части апекса периферическая меристема имеет ширину около 3 слоев клеток, а к основанию апекса она расширяется до 6–7 слоев. В этом направлении в ней возрастает концентрация нуклеопротеидов, митотическая активность клеток, конденсация хроматина ядер. Клетки периферической меристемы обладают минимальной вакуолизацией. Периферическая меристема представляет собой сложно организованную ткань. Гистохимическое изучение периферической меристемы показывает, что в ней существуют группы клеток с высоким содержанием нуклеопротеидов, которые участвуют в формировании зачатков хвои. Клетки одного и того же сектора периферической меристемы значительно варьируют по парциальному объему ядер, содержанию крахмала и гиалоплазмы. В клетках с большим объемом ядер объем гиалоплазмы минимальный. Группы клеток, содержащие повышенное количество сферосом, менее вакуолизированы и накапливают больше крахмала. Рис. 22. Вид сверху на апекс и зачаток охвоенного побега ели сибирской (Picea obovata) в период заложения фитомеров хвои (начало августа) в терминальной почке главной оси дерева. 1 – апекс, 2 – боковая поверхность зачатка охвоенной части побега с листовыми бугорками, 3 – листовые бугорки, расположенные на одной листовой спирали 48 Клетки самых молодых зачатков хвои по средним морфометрическим показателям ультраструктуры близки к периферической меристеме. Они имеют четкую тенденцию изменчивости показателей структуры в направлении от периферии к оси зачатка. В целом периферическую меристему апекса следует рассматривать как совокупность материнских групп клеток зачатков фитомеров (фитомер имеет в своем составе лист или другой боковой орган и принадлежащие ему элементы узла и междоузлия стебля). Они начинают свое формирование в виде относительно самостоятельных клеточных комплексов на границе зоны центральных материнских клеток и продолжают его в пределах периферической меристемы. Развитие материнских групп клеток зачатков фитомеров в периферической меристеме приводит к вычленению из нее в основании апекса боковых выростов – зачатков кроющих чешуй почки или листовых бугорков, называемых примордиями. В последующем из них развиваются кроющие чешуи почки, или листья. Наряду с этим, из клеток периферической меристемы происходит формирование меристематических клеток тканей проводящего цилиндра и коры почки и стебля зачатка побега. Каждый листовой бугорок первоначально развивается в виде полусферического выступа на поверхности в основании апекса. В его осевой части формируется прокамбиальный пучок, который связан с системой прокамбиальных пучков центрального цилиндра зачатка побега. Листовые бугорки появляются в основании апекса упорядоченно на нескольких листовых спиралях (парастихах), примыкающих друг к другу. Примордии хвои имеют при этом строго одинаковый размер и располагаются в правильном геометрическом порядке (рис. 22). Число листовых спиралей варьирует в зависимости от величины закладываемого зачатка побега. У наиболее крупного зачатка побега число парастих листовых примордиев в направлениях по часовой и против часовой стрелки достигает 21+34. По мере уменьшения размеров закладываемых зачатков побегов, количество парастих листовых бугорков изменяется в соответствии с рядом Фибоначчи, т.е. (13 + 21), (8 + 13), (5 + 8). Апекс и зачаток побега обеспечиваются продуктами жизнедеятельности главным образом через прокамбиальные пучки из расположенных ниже тканей зрелой части почки. Меристемы сердцевины и коровой паренхимы в конце вегетационного периода отделяются специализированными клетками диафрагмы от тканей почки. Жизнедеятельность апикальных меристем побегов хвойных осуществляется постоянно под покровами кроющих чешуй почки. В начале вегетационного периода апекс находится под покровами почки, которая сформировалась в предшествующий вегетационный период, а после ее распускания – под покровами почки новой генерации. 49 В апексе на протяжении всего вегетационного периода формируются материнские группы клеток зачатков различных типов фитомеров. Элементы новой почки начинают закладываться апексом весной в начале вегетационного периода, когда он в виде простого бугорка располагается на верхушке молодого побега. Этот зачаток побега, в свою очередь, еще находится под покровами набухающей почки предшествующей генерации. Заложение новой молодой почки происходит в продолжение гистологического развития и активного роста фитомеров охвоенной части молодого побега генерации предыдущего года. Развитие материнских групп клеток фитомеров почки характеризуется коротким периодом их формирования в апексе. Зачатки кроющих чешуй почки появляются в виде валикообразных бугорков в основании апекса. Они вытянуты вдоль периметра основания апекса. Примордии относительно быстро разрастаются и дифференцируются в кроющие чешуи почки – катафиллы. В начале периода заложения фитомеров почки апекс имеет относительно небольшие размеры и уплощенную форму верхушки. По мере уменьшения темпа продольного роста стебля охвоенной части материнского побега происходит замедление вычленения зачатков фитомеров почки. Это связано с затуханием митозов в основной паренхиме листьев и стебля и сохранением их активности в области камбия и феллогена. В апексе при этом начинается интенсивное заложение материнских групп клеток листовых фитомеров будущего зачатка охвоенной части побега. В результате прохождения этого процесса в конце этапа формирования почки (конец июня – начало июля) в апексе происходит накопление материнских групп клеток листовых фитомеров. Это приводит к значительному увеличению объема апекса (рис. 23). В вегетативных почках он приобретает правильную коническую форму. Такая форма апекса возникает вследствие равномерного разрастания слоев материнских групп клеток листовых фитомеров в периферической меристеме делением антиклинальными перегородками. В последующем каждая материнская группа клеток фитомера формирует в основании апекса полусферический листовой бугорок. Таким образом, из апекса начинают вычленяться зачатки фитомеров олиственной части побега. Они отличаются от фитомеров почки тем, что значительное время остаются в меристематическом состоянии, и в замедленном темпе проходят гистологическое развитие, накапливая в своем объеме значительное количество недифференцированных меристематических клеток. Данный характер развития материнских групп клеток фитомеров охвоенной части побега коррелирует с сохранением клеточных делений лишь в зонах вторичных меристем – камбия и феллогена стебля материнского побега. 50 1 2 33 4 55 6 Рис. 23. Вегетативная почка ели сибирской (Picea obovata) в конце этапа формирования почки: 1 – кроющая чешуя, 2 – верхушка апекса с апикальными инициалями и центральными материнскими клетками, 3 – периферическая меристема в основании апекса, 4 – сердцевинная меристема, 5 – проводящий пучок стеблевой части почки, 6 – сердцевина стеблевой части почки Во второй половине вегетационного периода темп вычленения зачатков фитомеров охвоенной части, побега начинает постепенно превышать темп заложения их материнских групп клеток в апексе. Это приводит к значительному уменьшению объема апекса в конце вегетационного периода (рис. 103). 51 Апексы побегов покрытосеменных растений У покрытосеменных растений клетки в самой верхней дистальной части апекса расположены слоями. Считается, что у этих растений в меристеме присутствуют два или несколько рядов клеток инициалей, расположенных один над другим. Морфологически инициали обычно неотличимы от первого поколения их дочерних клеток. Представление об обособленных рядах инициалей вылилось в концепцию организации апекса по типу туники-корпуса, предложенную в 1924 г. А. Шмидтом. Согласно этой теории, инициальная зона апикальной меристемы содержит две гистологические зоны: 1) тунику, включающую один или несколько периферических слоев клеток, которые делятся в плоскостях, перпендикулярных поверхности меристемы (антиклинальные деления); 2) корпус, состоящий из нескольких слоев клеток, делящихся в различных направлениях. Корпус представляет собой центральную группу клеток, в результате роста которых увеличивается объем апикальной меристемы. Туника представляет собой покров из одного или нескольких слоев клеток. Она разрастается над корпусом путем роста в поверхность. Число слоев туники у покрытосеменных варьирует. У двудольных в тунике насчитывается от 2-х до 4-5-и слоев клеток (рис. 24). У однодольных растений туника обычно содержит один или два слоя клеток. Рис. 24. Верхушка побега гороха (Pisum). А. Клеточное строение. Б. Схема направлений роста. 1 – антиклинальное деление; 2 – инициальный слой корпуса; 3 – двухслойная туника; 4 – листовой примордий; 5 – корпус; 6 – сердцевинная меристема 52 Дифференциация различных структур покрытосеменного растения, формирующихся из апекса, не предопределена конкретными слоями его клеток. Однако эпидерма всегда происходит из наружного слоя меристемы. Клетки второго слоя апекса образуют субэпидермальную часть периферической зоны. А клетки третьего слоя формируют центральную и периферическую зоны. В процессе заложения листового бугорка у однодольных растений принимает участие наружный слой апекса путем периклинальных делений клеток перегородками, параллельными внешней поверхности. При многослойной тунике весь листовой примордий может формироваться из ее производных. Интервал времени между закладкой двух последовательных листьев в апексе принято обозначать пластохроном. Маргинальная меристема Листовые бугорки образуются в форме боковых выступов апекса. Ранний рост листового бугорка – примордия подразделяется на апикальный и маргинальный. Первоначально примордий разрастается в длину за счет деятельности его апикальной меристемы. Дальнейшее формирование плоской листовой пластинки из такого бугорка осуществляется деятельностью маргинальной меристемы (рис. 25). Она образуется из двух валиков меристематических клеток, расположенных по краям бокового выступа. Каждый такой валик представляет собой маргинальную меристему. У двудольных растений наружный слой клеток маргинальной меристемы делится антиклинальными перегородками, перпендикулярными поверхности листового зачатка. Таким образом осуществляется увеличение площади покровной ткани листа. Второй слой клеток маргинальной меристемы в основном обеспечивает формирование слоев ассимиляционной паренхимы листовой пластинки, а также прокамбиальных пучков, из которых развиваются проводящие пучки. Наряду с деятельностью маргинальной меристемы рост листовой пластинки также осуществляется за счет продолжающихся делений клеток, расположенных в основании развивающегося листа. 53 Рис. 25. Развитие листовой пластинки двудольного растения (Nicotiana tabacum): I – III. Последовательные стадии роста бокового края листовой пластинки на поперечном срезе. Маргинальная меристема состоит из наружных маргинальных инициалей (1), и субмаргинальных инициалей (3,5). Антиклинальные деления производных маргинальных инициалей (2) приводят к формированию нижней и верхней эпидермы листа. Антиклинальные деления субмаргинальных инициалей (3,5) обеспечивают формирование клеток ассимиляционной паренхимы – палисадной (4) и губчатой (7). Периклинальные деления субмаргинальных инициалей служат источником заложения прокамбиальных пучков (8), из которых развиваются проводящие ткани листа в виде ксилемы (6) и флоэмы. IV. Продольный разрез верхушки листового зачатка в период первичного роста в длину. Апикальная инициаль (9) производит клетки верхней и нижней эпидермы. Субапикальная инициаль (10) антиклинальными делениями инициирует слои мезофилла, а в результате периклинальных делений формируются клетки (11), из которых развивается прокамбий (8) АПИКАЛЬНАЯ МЕРИСТЕМА КОРНЯ Формирование первого корня семенного растения начинается с развития его апикальной меристемы на нижнем конце гипокотиля зародыша семени. Эта апикальная меристема в последующем при развитии растения формирует его стержневой корень (первичный корень). Главный корень и формируемые им боковые корни образуют стержневую корневую систему, проникающую глубоко в почву. Такая система характерна для голосеменных и двудольных растений. У однодольных растений первичный корень функционирует сравнительно недолго и заменяется несколькими придаточными корнями, возникающими на побеге совместно с пазушными почками. Эти корни образуют относительно однородную мочковатую корневую систему, которая хорошо закрепляет растение в поверхностных слоях почвы. 54 Положение слоев меристематических клеток в апексе корня довольно часто предопределяет характер формируемых им тканей. Это послужило основанием для разработки во второй половине XIX в. Дж. Ганштейном теории гистогенов. Согласно этой теории в апикальной меристеме существуют слои клеток гистогены, из которых развивается тело растения. Каждый гистоген берет начало от одной или нескольких инициалей, расположенных ярусами друг над другом. Различаются следующие гистогены: 1) дерматоген, являющийся предшественником эпидермы; 2) периблема, из которой формируется первичная кора; 3) плерома, дающая начало центральному цилиндру. Подразделение апекса на три гистогена редко выявляется в апикальных меристемах побегов и репродуктивных органов, но в некоторых случаях оно оправдано для описания дифференциации тканевых зон в корне. Наряду с этим, для характеристики взаимоотношений между характером расположения, роста и деления клеток, а также ростом корня, в начале XX века разработана теория тела-чехлика КорпераКаппе. Данная теория учитывает направление плоскостей делений клеток, ответственных за увеличение числа продольных клеточных рядов в меристематической зоне корня. В частности, при клеточном росте продольные и поперечные деления распределяются таким образом, что в месте раздвоения рядов клеток поперечные и продольные стенки в данном ряду располагаются в виде буквы т (или y). Если корневой чехлик имеет собственные инициали, то направление, соответствующее горизонтальной перекладине буквы т в клеточных рядах, резко отделяет корневой чехлик от тела корня. Линия с изломом, соответствующая горизонтальной перекладине буквы т, в корневом чехлике обращена к основанию корня, а в теле Рис. 26. Схема строения апекса корня папоротника корня – к его апексу (рис. 26). Дан(Dennstaedtia): ная теория также дает представле1 – апикальная инициальная ние о минимальном числе клеток в клетка, 2 – ризодерма, апексе, которое необходимо для 3 – эндодерма, 4 – центральный создания при их делении характерцилиндр, 5 – корневой чехлик; ной структуры корня (например, одТ – Т-образная перекладина в клеточных рядах в теле корня на у многих низших растений). 55 Подробные физиологические и биохимические исследования роста корня показали, что увеличение числа клеток в его кончике, вероятно, не зависит от непрерывной активности наиболее удаленной небольшой группы апикальных клеток. Инициали, ответственные за организацию клеточного роста на первых стадиях роста корня, Рис. 27. Схема распределения митотической активности на продольном разрезе в кончика корня лука (Allium). Частота митозов соответствует плотности точек. 1 – корневой чехлик; 2 – эпидерма; 3 – первичная кора; 4 – ситовидный элемент протофлоэмы; 5 – центральный цилиндр; 6 – покоящийся центр в основном утрачивают митотическую активность на последующих стадиях роста корня. При этом более высокую митотическую активность проявляют клетки, расположенные на некотором удалении от верхушки корня, где находятся инициали (рис. 27). Данные исследования послужили основой для формулирования Ф. Клаусом в 60-х гг. XX в. теории покоящегося центра в апикальной меристеме. В соответствии с этой теорией самые удаленные клетки в теле корня – инициали плеромы и периблемы, делятся редко, имеют приблизительно одинаковые размеры и низкие скорости синтеза нуклеиновых кислот и белков. Применение методики включения в ядра клеток 3Нтимидина, меченного радиоактивным тритием, и блокирования клеточного деления в метафазе ингибиторами позволило получить количественные данные о продолжительности митотического цикла в различных зонах верхушки корня. На основании этих исследований установлено, что клетки покоящегося центра делятся приблизительно в десять раз медленнее, чем соседние клетки меристемы. Считается, что покой у клеток корневой меристемы наступает вследствие антагонистической направленности клеточного роста в различных частях меристемы, причем особенно важную роль в подавлении роста играет корневой чехлик. В соответствии с другим 56 мнением, причиной отсутствия активности в самом покоящемся центре является наличие быстро делящихся вокруг него клеток. В том случае, когда центральный цилиндр, первичная кора и корневой чехлик имеют собственные инициали, а эпидерма дифференцируется из наружного слоя первичной коры или из клеток, относящихся к корневому чехлику, апикальную меристему относят к закрытому типу. А когда все гистологические зоны корня развиваются из одной группы инициалей, такую апикальную меристему относят к открытому типу. Считается, что открытый тип апикальной меристемы является филогенетически более примитивным, чем закрытый тип. По мере роста молодого корня апикальная меристема приобретает определенную структурную организацию, которая имеет характерные особенности для разных таксонов растений. Наиболее просто устроен апекс корня сосудистых споровых растений, таких как папоротники, хвощи, плауны. У этих таксонов апекс имеет единственную апикальную инициаль, из которой формируются все ткани корня (рис. 26). Апекс находится под покровами корневого чехлика, который в виде колпачка предохраняет меристематические клетки от повреждения при продвижении растущего корня между частицами почвы. Формирование тканей корня из единственной апикальной инициальной клетки апекса осуществляется следующим образом. Вперед по оси корня вычленяются меристематические клетки корневого чехлика. В стороны вычленяются меристематические клетки ризодермы и первичной коры, которые располагаются под корневым чехликом. А по оси в направлении зрелых проводящих тканей апикальная инициаль формирует меристематические клетки центрального цилиндра. Более сложно устроен апекс корня у голосеменных и покрытосеменных растений. В частности, у голосеменных одна группа инициальных клеток апекса формирует меристематические клетки ризодермы, первичной коры и проводящего цилиндра. Эта же группа инициалей образует колонку меристематических клеток, из которой развиваются слои корневого чехлика (рис. 28, Д,Е). У покрытосеменных двудольных и однодольных растений апекс корня содержит три яруса инициальных клеток (рис. 28 А, Б, В, Г). Из верхнего яруса инициалей формируется проводящий цилиндр. Из среднего яруса инициалей образуются меристематические клетки, формирующие ризодерму и первичную кору. Нижний ярус инициалей образует калиптроген, дающий начало корневому чехлику. 57 Рис. 28. Апикальная меристема корня и продуцируемые ею гистологические зоны. А, Б. Три яруса инициалей корня однодольного растения (ковыль – Stipa), включающих инициали чехлика, инициали коры и ризодермы, а также инициали центрального цилиндра. В, Г. Три яруса инициалей корня двудольного растения (редис – Raphanus). Из нижнего яруса формируются ризодерма и корневой чехлик; на рисунке (Г) стрелками показана его внутренняя граница). Средний ярус продуцирует клетки коры, а верхний – клетки центрального цилиндра. Д–Е. Апекс голосеменного растения (ель – Picea). Все зоны корня происходят из одной группы инициалей. Из этой группы формируются центральный цилиндр, кора, ризодерма, а также колонка меристематических клеток корневого чехлика (колумела). От этой колонки в латеральном направлении возникают производные клетки, из которых форимруется чехлик. 1 – центральный цилиндр; 2 – первичная кора; 3 – перицикл; 4 – слизистый слой; 5 – ризодерма; 6 – калиптроген; 7 – корневой чехлик; 8 – колонка 58 Заложение бокового корня Формирование нового бокового корня у большинства цветковых растений начинается на значительном удалении от апикальной меристемы материнского корня. В то же время у некоторых видов, например у представителей тыквенных (сем. Cucurbitaceae) и ряда водных растений, примордии боковых корней закладываются вблизи от кончика корня, среди клеток, которые не утратили способность делиться. Процесс инициации боковых корней, например у проростков пшеницы (Triticum aestivum), начинается с возобновления деления клеток в перицикле на значительном удалении – 16–18 мм от кончика корня и его меристемы. Клетки перицикла в данной зоне прозенхимные, имеют длину 250–300 мкм. Здесь также возобновляется деление клеток стеллярной паренхимы. На этом уровне корня наблюдается значительная дифференциация элементов ксилемы: клетки протоксилемы уже безъядерные, а у клеток периферической метаксилемы сформировались утолщенные оболочки (рис. 29, IV). Деление клеток перицикла происходит только перегородками, поперечными продольной оси корня, в разных продольных рядах по периметру центрального цилиндра корня. При этом в секторе между двумя продольными рядами протоксилемы, где осуществляется инициация примордия бокового корня, у клеток перицикла обнаруживается более тонкая клеточная оболочка, чем в остальных секторах. Здесь, как правило, происходят неравные деления клеток перицикла, приводящие к образованию длинной и короткой дочерних клеток. В секторе инициации примордия бокового корня первыми осуществляют неравные деления клетки двух продольных рядов перицикла, расположенные против ситовидной трубки флоэмы, перегородками, поперечными этим рядам. В результате таких делений в каждом продольном ряду перицикла образуются пара коротких (длиной 50–80 мкм) и две, по сторонам от них, длинные (130 мкм и более). После первых делений клеток перицикла в секторе инициации нового бокового корня начинается пролиферация клеток эндодермы. Деление клеток эндодермы в секторе инициации примордия бокового корня также происходит перегородками, поперечными продольной оси материнского корня. Они бывают равными и неравными. В инициации примордия бокового корня обычно принимают участие 8–12 клеток материнского корня. После завершения одного-двух поперечных неравных делений клеток перицикла начинается их радиальный рост в направлении 59 поверхности корня. Первыми начинают радиальный рост самые короткие клетки перицикла, расположенные в двух соседних рядах, контактирующих с ситовидными элементами протофлоэмы. Эти клетки раньше, чем сестринские им клетки, начинают периклинальные деления. В конечном итоге базальная клетка перицикла, контактирующая с ситовидным элементом протофлоэмы, оказывается большей по размеру и имеет более крупную вакуоль, чем сестринская апикальная клетка. В последующем митотическом цикле базальная клетка разделяется раньше, чем апикальная. В результате дальнейшего роста и деления клеток перицикла антиклинальными и периклинальными перегородками формируется примордий бокового корня, окруженный кармашком, состоящим из клеток эндодермы. Этот кармашек образуется за счет антиклинальных (поперечных и радиальных) делений клеток в трех-четырех продольных рядах эндодермы, расположенных против сектора инициации примордия бокового корня в перицикле. Наряду с этим, возобновляется деление и некоторых клеток коры перегородками, поперечными продольной оси материнского корня. Эти клетки не участвуют в формировании кармашка. Клетки в основании эндодермального кармашка, по мере роста примордия бокового корня, прекращают рост и деление, а затем вакуолизируются. При этом расположенные на флангах примордия клетки перицикла разрастаются, сминают производные клеток эндодермы и разрывают кармашек. В верхней части примордия клетки эндодермы продолжают рост и деление. Перед выходом примордия бокового корня на поверхность материнского корня клетки кармашка образуют эндодермальный чехлик. К этому времени в примордии формируются инициальные клетки чехлика, ризодермы и коры перициклического происхождения (рис. 30, II). После первых делений клеток стеллярной паренхимы перегородками, поперечными продольной оси корня, начинается их деление периклинальными и антиклинальными радиальными перегородками. Это приводит к разрастанию стеллярной паренхимы, расположенной в основании примордия бокового корня, в радиальном и тангенциальном направлениях. В результате такого роста происходит увеличение диаметра основания примордия и расстояния между его основанием и центральным рядом клеток метаксилемы, а также числа клеток стеллярной паренхимы материн60 Рис. 29. Инициация и начальные этапы развития примордиев боковых корней в первичном корне Triticum aestivum. I – продольный срез кончика первичного корня, общий вид. II, III – продольные срезы первичного корня на удалении 18–20 мм от его кончика. Митоз в центре длинной клетки перицикла, приводящий к равному делению (II), завершены антиклинальные неравные деления в двух соседних клетках перицикла (III): IV, V – поперечные срезы в секторе инициации примордия на удалении 24 и 26 мм от кончика корня. Стрелками отмечен сектор инициации примордия. (IV – оболочки клеток перицикла в секторе закладки примордия более тонкие, чем в остальных секторах. V – начало радиального роста клеток перицикла в секторе инициации примордия). VI – продольный срез материнского корня на удалении 30 мм от его кончика. В перицикле завершены два первых периклинальных деления клеток, а в эндодерме – неравные деления двух соседних в ряду клеток. 1 – кора, 2 – эндодерма, 3 – протоксилема, 4 – перицикл, 5 – метаксилема, 6 – флоэмная группа, 7 – стелярная паренхима, 8 – осевой ряд метаксилемы, 9 – чехлик. Масштабная линейка: I – 150 мкм; II, III, IV, V, VI – 25 мкм 61 ского корня в основании развивающегося примордия бокового корня (рис. 30, I, II). В процессе продвижения на поверхность примордий бокового корня сминает клетки коры материнского корня (рис. 30, I, II). Затем растущий примордий прорывает экзодерму и ризодерму материнского корня и выходит на его поверхность (рис. 30, III), что происходит обычно на удалении 70–90 мм от кончика материнского корня. При этом боковой корень остается окруженным кармашком, клетки которого в апикальной части еще остаются живыми. Чехлик бокового корня увеличивается в размерах, но колумела (колонка в центральной части корневого чехлика) еще не обнаруживается. После выхода примордия бокового корня на поверхность материнского корня начинается этап активизации деятельности инициальных клеток разных тканей бокового корня. За счет роста и деления инициалей и их производных формируются продольные ряды клеток. В боковом корне длиной 0,5–0,6 мм уже выявляется меристема, а в базальной части начинается растяжение клеток (рис. 30, III). В дальнейшем, по достижении боковым корнем длины 1,0–1,5 мм и диаметра 150–200 мкм, в нем осуществляется формирование ростовых зон: меристемы и зоны растяжения. Клетки корневого чехлика начинают выделение полисахаридной слизи. Длина меристемы в боковом корне значительно меньше, чем в материнском. В боковом корне начинается дифференциация проводящей системы и зоны контакта с проводящей системой материнского корня. В зоне контакта дифференцируются клетки стеллярной паренхимы. В клетках, контактирующих с флоэмной группой, осуществляется дегенерация ядер, и эти клетки дифференцируются в ситовидные элементы с живым протопластом. Клетки, контактирующие с периферической метаксилемой, дифференцируются в трахеальные элементы. В них формируются вторичные утолщения клеточных оболочек. В результате дифференциации клеток стеллярной паренхимы число проводящих элементов ксилемы и флоэмы в секторе закладки бокового корня в материнском корне увеличивается. Далее процессы дифференциации ситовидных и трахеальных элементов распространяются в боковой корень. Формируется проводящая система бокового корня с четырьмя-пятью флоэмными и ксилемными тяжами. 62 Рис. 30. Развитие примордиев боковых корней в первичном корне проростков Triticum aestivum. I, II – поперечные срезы первичного корня на удалении 40–60 мм от его кончика. Примордий на ранних стадиях развития в материнском корне. Завершились периклинальные деления в клетках стелярной паренхимы (отмечено стрелками). (I – примордий окружен эндодермальным кармашком; II – непрерывность кармашка нарушена, инициальная группа клеток перициклического происхождения находится в примордии). III, IV – продольные срезы первичного корня на удалении 80 и 100 мм от его кончика. III – начало формирования зоны растяжения бокового корня. IV – зона контакта проводящих систем материнского и бокового корней. 1 – кора, 2 – эндодерма, 3 – протоксилема, 4 – перицикл, 5 – метаксилема, 6 – флоэмная группа, 7 – стелярная паренхима, 8 – осевой ряд метаксилемы, 9 – чехлик, 10 – эндодермальный чехлик, 11 – инициальные клетки чехлика, 12 – зона растяжения, 13 – трахеальные элементы, соединяющие ксилему материнского и бокового корней, 14 – меристема, 15 – инициальная клетка ризодермы и коры, 16 – инициальные клетки центрального цилиндра. Масштабная линейка: I, II, IV – 25 мкм, III – 50 мкм 63 ЛАТЕРАЛЬНЫЕ МЕРИСТЕМЫ Первичные латеральные меристемы Прокамбий Прокамбий представляет собой первичную латеральную образовательную ткань, которая развивается в зачатках органов из клеток периферической меристемы апекса, а также из клеток, расположенных между нею и сердцевинной меристемой (рис. 21). Он образует пучки, которые группируются в виде центрального сетчатого цилиндра, с ответвлениями в листовые примордии. Прокамбий состоит из молодых, относительно вакуолизированных клеток. Они имеют повышенное содержание нуклеиновых кислот, обладают высоким метаболизмом, митотически активны и выполняют функцию транспорта веществ между проводящим цилиндром стеблевой части почки, зачатком побега и апексом. Весной, когда активизируется рост зачатка побега в почке, клетки прокамбия дифференцируются в проводящие ткани – первичную флоэму и первичную ксилему. В первую очередь дифференцируются элементы флоэмы. Она формируется с внешней стороны от прокамбия. Несколько позже созревают первые проводящие элементы ксилемы. В период начального роста органов, когда увеличиваются их продольные и поперечные размеры, прокамбий формирует достаточно эластичные элементы протофлоэмы и протоксилемы. После завершения продольного роста органов из клеток прокамбия развиваются элементы метафлоэмы и метаксилемы, имеющие наибольшие размеры и более жесткие клеточные оболочки. Характерная особенность строения первичной флоэмы и первичной ксилемы состоит в том, что их клетки располагаются относительно произвольно без заметной системы взаимного расположения. У растений, обладающих способностью к вторичному росту – голосеменных и двудольных, не все клетки прокамбия дифференцируются в постоянные проводящие ткани. Часть из них сохраняет митотическую активность и переходит к выполнению функций вторичной латеральной меристемы – камбия. В молодом корне по оси формируется один прокамбиальный пучок. Из этого пучка развивается проводящий пучок радиального типа. Он состоит из центрально расположенного ребристого цилиндра первичной ксилемы, во впадинах которой с внешней стороны лежат отдельные тяжи первичной флоэмы. У однодольных 64 растений прокамбий в корне функционирует ограниченное время, и его клетки полностью дифференцируются в проводящие ткани. У корней голосеменных и двудольных растений между первичной флоэмой и первичной ксилемой остаются молодые меристематические клетки, которые дают начало вторичной латеральной меристеме – камбию. Перицикл Перицикл относится к первичной латеральной меристеме. Он представляет собой слой клеток, расположенный, как правило, в корне под эндодермой – внутренним слоем первичной коры. Часть клеток перицикла сохраняет меристематическую активность и участвует в формировании апексов боковых корней (рис. 29, 30). Кроме этого у голосеменных и двудольных растений из клеток перицикла закладывается феллоген – пробковый камбий, а также частично и камбий. Вторичные латеральные меристемы Камбий Камбий представляет собой вторичную боковую меристему, функционирующую обычно на протяжении всей жизни многолетних органов растения. В корне и стебле он формирует молодые клетки вторичной флоэмы и вторичной ксилемы. Характерная особенность деятельности камбия состоит в том, что продуцируемые им клетки флоэмы и ксилемы располагаются в виде правильных радиальных рядов. Это связано с тем, что камбий имеет один слой инициальных клеток, расположенных в виде цилиндра. Каждая инициальная клетка периодически вычленяет, в результате деления периклинальной перегородкой, во внешнюю сторону материнскую клетку флоэмы, а затем в направлении к оси органа – материнскую клетку ксилемы (рис. 31). Причем, в сторону ксилемы откладывается большее число молодых клеток, чем в сторону флоэмы. Это приводит к преобладанию нарастания объема древесины над лубом. Молодая меристематическая клетка после вычленения от инициальной клетки камбия делится также периклинально один или несколько раз, а затем дифференцируется в клетки флоэмы или ксилемы. Таким образом, слой инициальных клеток камбия, а также ближайших производных от них флоэмных и ксилемных инициалей, сохраняющих меристематическую активность, образуют камбиальную зону. 65 Рис. 31. Камбий и формируемые им ткани. Веретеновидная (А) и лучевая (В) инициали камбия. С, D – вторичная флоэма, камбий и вторичная ксилема Robinia pseudoacacia. C. Поперечный срез. D. Радиальный срез (D-I – элементы осевой системы, D-II – элементы радиального луча). 1 – осевые инициали; 2 – периклинальная плоскость деления инициалей, в результате которого образуются новые клетки флоэмы и ксилемы; 3 – лучевые инициали; 4 – вторичная ксилема; 5 – крупный сосуд; 6 – сосуды меньшего размера; 7 – камбий; 8 – вторичная флоэма; 9 – ситовидные трубки; 10 – лубяные волокна Камбий имеет вытянутые веретеновидные инициальные клетки, из которых развиваются удлиненные элементы флоэмы и ксилемы. Относительно короткие лучевые инициальные клетки дают начало радиальным слоям паренхимных клеток. Хвойные растения, более примитивные в эволюционном отношении, чем покрытосеменные, образуют большие клетки веретеновидных инициалей, имеющих длину до 3–4 мм. Среди покрытосеменных у менее эволюционно продвинутых двудольных длина этих клеток составляет 0,8–,3 мм, а у более продвинутых – 0,15–0,17 мм. Веретеновидные инициали с возрастом растения увеличиваются по длине. Например, у однолетней сосны длина веретеновидных инициалей камбия составляет 0,87 мм, а у 60-летнего экземпляра 66 достигает 4,0 мм. При дальнейшем увеличении возраста длина этих клеток существенно не изменяется. Веретеновидные инициальные клетки образуют все продольно вытянутые элементы луба и ксилемы – ситовидные трубки, лубяные волокна, осевую лубяную паренхиму, сосуды, трахеиды, волокна либриформа и осевую древесную паренхиму. Лучевые инициальные клетки собраны в группы, с линзовидными очертаниями на тангенциальном сечении. Эти клетки имеют меньшие продольные размеры, чем веретеновидные инициали. Каждая лучевая инициаль продуцирует радиальный ряд паренхимных клеток как в сторону флоэмы, так и в сторону ксилемы. В связи с этим, лучи следуют в этих тканях относительно прямолинейно, не прерываясь в камбии. Во время деления веретеновидной инициали продольной перегородкой фрагмопласт, образующийся во время телофазы, намного короче, чем длинная ось клетки (рис. 32). При этом вскоре после деления ядра фрагмопласт и клеточная пластинка достигают продольных стенок клетки. Дальнейшее продвижение клеточной пластинки в направлении длинной оси к противоположным концам клетки происходит в течение более длительного времени. Активизация камбия у древесных растений умеренных широт после зимнего периода покоя наступает весной. При этом его клетки поглощают воду, растягиваются в радиальном направлении и делятся тангенциальными перегородками. Радиальные стенки камбиальных клеток становятся более тонкими, что приводит к уменьшению прочности связи коры со стеблем. В результате этого кора начинает легко отделяться от древесины, что может служить признаком весенней активизации камбия. Первые деления в камбиальной зоне весной происходят в перезимовавших материнских клетках флоэмы и ксилемы. И лишь затем начинается митотическая активность непосредственно в инициальном слое камбия. У покрытосеменных растений, например Eucaliptus, отложение клеток в сторону ксилемы больше в четыре раза, чем в сторону флоэмы. Еще более значительное преобладание отложения ксилемы над флоэмой обнаружено у хвойных растений, например у Tuja occidentalis. В период интенсивного роста у этого растения на флоэмную сторону откладывается 12–16 клеток, а на ксилемную – 100 и более. Возобновление камбиальной активности весной тесно взаимосвязано с возобновлением роста зачатков побегов в почках. В первую очередь камбиальная активность появляется в стеблях зрелых годичных побегов, несущих почки с зачатками побегов. Затем она 67 Рис. 32. Деление веретеновидных инициалей камбия тангенциальной перегородкой. I–III – завершение деления ядра (кариокинез) и три стадии деления клетки (цитокинез) (радиальные срезы). IV–V – две стадии формирования клеточной пластинки (тангенциальные срезы). 1 – фрагмопласт; 2 – клеточная пластинка; 3 – ядра; 4 – новая клеточная оболочка между двумя дочерними клетками распространяется к основанию ветвей в ствол и далее по нему к корням. Таким образом, рост зачатков побегов в набухающих почках весной стимулирует возобновление ростовых процессов в расположенных ниже осях. Инициация камбиальной активности связана с активизацией синтеза ростовых веществ, прежде всего ауксинов, в развивающихся почках. Ауксины транспортируются в нисходящем направлении по флоэме. Поддержание дальнейшей камбиальной активности осуществляется затем дифференцирующимися клетками ксилемы. Это связано с тем, что ауксин начинает синтезироваться в процессе дифференциации и автолиза трахеальных клеток, которые превращаются в мертвые проводящие элементы. Наряду с ауксином камбиальную активность стимулируют также другие фитогормоны, например, цитокинины и гибберелловая кислота, которые совместно могут активизировать деление камбиальных клеток и дифференциацию их производных. 68 Гормональные регуляторы обычно действуют в сочетании с другими ростовыми факторами, к которым относятся питательные вещества, особенно сахара, вода, благоприятная температура, оптимальный фотопериод, а также собственные эндогенные ритмы ростовых процессов в растении. Первые деления камбиальных клеток в ветвях многолетних растений в умеренных широтах отмечаются в конце апреля, а первые деления в корнях – в начале июля. Осеннее прекращение активности камбия первоначально наступает в ветвях в конце июля, а в корнях – значительно позже, в конце сентября. Камбиальная деятельность может прекращаться также в продолжение вегетационного периода в результате уничтожения листвы насекомыми, резкой засухи и т.д. В тропических областях деятельность камбия прерывается в сухие периоды года, а в районах, где сухие периоды отсутствуют или выражены слабо, камбий функционирует относительно равномерно в течение всего года. Рис. 33. Типы структуры камбия по расположению веретеновидных и лучевых инициалей. А. Беспорядочное распределение камбиальных клеток. Б. Частичная ярусность камбия. В. Полная ярусность камбия Однако и здесь также возможно замедление и полная остановка работы камбия на некоторый срок. По мере увеличения окружности цилиндра камбия, что связано с общим ростом органа, камбиальные инициали продуцируют свои производные также и в тангенциальном направлении. Эти новые инициали начинают формировать дополнительные радиальные ряды клеток вторичной флоэмы и ксилемы. В зависимости от расположения веретеновидных и паренхимных инициалей в тангенциальной плоскости (параллельной наружной 69 поверхности органа) различают несколько типов строения камбия (рис. 33). При отсутствии системы во взаимном расположении веретеновидных и лучевых инициалей их размещение приято обозначать в камбии как беспорядочное. В том случае, когда веретеновидные инициали образуют правильные тангенциальные ряды, а лучевые располагаются произвольно между ними, размещение клеток камбия считается как частично ярусное. Правильное размещение веретеновидных и лучевых инициалей в виде тангенциальных рядов создает полную ярусность камбия. Феллоген Феллоген представляет собой вторичную латеральную меристему. Она формирует вторичную покровную ткань перидерму, состоящую из пробки, феллогена и феллодермы. Первоначально клетки феллогена у надземных органов растения чаще всего развиваются из живых паренхимных клеток, расположенных под первичной покровной тканью эпидермой, либо непосредственно из самой эпидермы (рис. 34). У корней феллоген закладывается в большинстве случаев в перицикле – первичной латеральной меристеме, окружающей прово- Рис. 34. Заложение феллогена в стебле пеларгонии и формирование им перидермы. I – феллоген закладывается в результате периклинальных делений субэпидермального слоя клеток первичной коры; II – завершение закладки перидермы. 1 – закладка феллогена, 2 – эпидерма, 3 – клетки пробки (феллемы), 4 – феллоген, 5 – феллодерма, 6 – тонкостенные молодые клетки первичной коры, 7 – зрелые клетки первичной коры с утолщенными оболочками 70 дящий цилиндр корня. Заложение феллогена связано с активизацией митотической деятельности живых клеток, которые, в основном, остаются потенциально меристематическими. Они не отличаются от других клеток того же типа. Обычно первый слой феллогена закладывается сразу по всей окружности оси, или отдельными участками, которые впоследствии соединяются в процессе латерального распространения меристематической активности. Феллоген закладывается в результате деления исходных клеток периклинальными перегородками. Такими же периклинальными делениями феллоген формирует во внешнюю сторону многочисленные слои молодых клеток пробки, расположенные в виде радиальных рядов. Эти клетки быстро созревают, их оболочки пропитываются суберином, пробковеют и становятся непроницаемыми для воды и газообразных веществ. В направлении к оси органа феллоген вычленяет единичные слои живых паренхимных клеток феллодермы. Обычно феллоген функционирует в продолжение одного вегетационного периода. В следующем году он вновь закладывается из паренхимных клеток, расположенных под прежним слоем перидермы. В результате такого наслоения перидерм образуется чешуйчатая или многослойная третичная покровная ткань – корка. ИНТЕРКАЛЯРНЫЕ МЕРИСТЕМЫ У однодольных и некоторых двудольных растений в основании зрелых частей листьев и междоузлий стебля остаются молодые клетки, обладающие меристематической активностью. Они относятся к интеркалярным меристемам и представляют собой зоны формирования новых молодых клеток (рис. 35). Рост и дифференциация этих клеток обеспечивает продольное удлинение Рис. 35. Схема расположения зрелых стебля и листа. Наличие интеркалярных меи меристематических ристем способствует одновременному росту тканей в стебле ржи нескольких междоузлий, что обусловливает, (Secale sereale): в конечном итоге, быстрый рост однодоль1 – интеркалярные меристемы; ных растений, например злаков. Зачатки ли2 – молодые формируюстьев покрытосеменных и хвойных растений щиеся ткани; также имеют в основании молодые ткани, 3 – зрелые ткани 71 клетки которых сохраняют способность к делению более продолжительное время, чем на верхушках. Поэтому данные зоны зачатков листьев также относят к интеркалярным меристемам. РАНЕВЫЕ МЕРИСТЕМЫ В местах повреждения органов растения на открытой поверхности оказываются разнообразные ткани. Самые внешние слои их клеток усыхают и погибают, выделяя при этом вещества, стимулирующие возобновление митотической активности у живых дифференцированных клеток близко расположенных тканей (рис. 36). Такую способность обычно приобретают тонкостенные паренхимные клетки. В результате дедифференциации (омоложения) живых клеток и активизации их деления под наружной раневой поверхностью нарастает новая паренхимная ткань – каллус (от лат. сallus – мозоль). Впоследствии клетки внешних слоев каллуса преобразуются в феллоген, который формирует на месте поранения новую перидерму. Рис. 36. Образование каллуса Каллусообразование широко на открытой поверхности ксилемы используется для вегетативного стебля гибискуса (Hibiscus). размножения растений привив1 – тонкостенные клетки каллуса, ками и черенками. При выполвозникшие из не полностью нении прививки почка или часть дифференцированных клеток побега прививаемого растения ксилемы – 2, в основном, из клеток лучевой паренхимы. помещается срезанной поверхВпоследствии под поверхностью ностью на надрез на стебле каллусных клеток возникает подвоя или в его расщепленную феллоген, формирующий часть. В процессе срастания новую перидерму, которая защищапрививки зоне контакта подвоя ет пораненный участок стебля. и привоя формируется каллусОдновременно с этим, глубже среди каллусных клеток ная ткань. Клетки ее специализакладывается камбий, который зируются в выполнении функначинает формировать вторичную ций близлежащих тканей, наксилему и вторичную флоэму пример, камбия и феллогена. над местом поранения стебля 72 Если поместить на 12-15 дней во влажную камеру при комнатной температуре отрезок стебля одревесневшего побега, то на его морфологически нижнем конце образуется большее накопление каллуса, чем на верхнем. Кроме того, вблизи каллусного нароста на нижнем конце черенка возникают придаточные корни. Эта полярность каллусообразования связана с повышением концентрации ауксинов, базипетально перемещающихся по флоэме и стимулирующих усиленное деление клеток. В данных условиях также происходит переключение морфогенетической программы клеток побега на формирование тканей корня. ПОКРОВНЫЕ ТКАНИ Покровные ткани окружают наружную поверхность органов растения. Они защищают внутренние ткани от излишней потери воды, механических и термических повреждений, свободного проникновения внутрь патогенных организмов и обеспечивают необходимый уровень обмена веществ между растением и внешней средой. Покровные ткани по времени образования подразделяются на первичную, к которой относится эпидерма, вторичную – перидерму и третичную – корку. Эпидерма Эпидерма возникает из протодермы – наружного слоя клеток меристематического апекса. Эпидерма окружает одним или несколькими слоями клеток органы растения. Для обозначения особых функций этой ткани у корней ее называют ризодермой или эпиблемой. Эпидерма имеет несколько типов клеток. Бόльшую часть этой ткани составляют неспециализированные основные эпидермальные клетки. Они выполняют, главным образом, защитные функции. Эти клетки в виде пластинок или плиток покрывают относительно ровным слоем поверхность органов. Поэтому их форму называют таблитчатой. У двудольных растений основные эпидермальные клетки листьев в периклинальной плоскости (параллельной наружной поверхности) преимущественно изодиаметрической формы. Удлиненные части органов у однодольных растений – стебли, черешки, листья, цветоносы, имеют эпидермальные клетки вытянутые в направлении длинной оси органа. 73 Основные клетки эпидермы – живые и содержат бесцветные пластиды – лейкопласты, способные запасать крахмал и белковые кристаллы. Вакуоли могут накапливать пигмент – антоциан, придающий окраску лепесткам цветков и плодам. В основных клетках эпидермы встречаются кристаллы. Наружная стенка клеток основной эпидермы, как правило, имеет наибольшую толщину. Ее целлюлозная оболочка снаружи покрыта тонкой прослойкой пектиновых веществ, удерживающих на поверхности слой жирового вещества – кутина значительной толщины, покрытого сверху воском (рис. 37). Кутиновый и восковой покровы наружных стенок эпидермальных клеток служат для уменьшения потери воды растением. В наружной оболочке эпидермальных клеток имеются тейходы или эктодесмы, представляющие собой узкие участки, в которых оболочка более рыхлая и проницаемая, чем Рис. 37. Схема организации в других местах. наружной оболочки клеток Кроме кутина в оболочках клеверхней эпидермы листа груши ток эпидермы накапливается лиг(Pirus communis) нин, вызывающий их одревеснес детальным изображением ние. Это характерно для листьев кутикулярного и воскового слоев: саговников, злаков, осок и иногда 1 – кутиновый матрикс, для двудольных – дубов, эвка2 – эпикутикулярный воск, липтов, лавра, олеандра. 3 – кутикула, 4 – пектиновые У хвощей, папоротников, вещества, 5 – целлюлозная обозлаков, осоковых, пальм и неколочка, 6 – двоякопреломляющий торых двудольных в оболочках слой воска, погруженный в кутин, клеток эпидермы откладывается 7 – антиклинальная стенка оболочки двух соседних клеток кремний. У представителей двудольных растений из семейств мальвовых, тутовых, горечавковых часть эпидермальных клеток ослизняется, что также характерно для эпидермы семян, например у льна. К специализированным клеткам эпидермы относятся замыкающие клетки устьиц (рис. 38). Рядом с ними у некоторых растений расположены побочные или околоустьичные клетки, которые принимают участие в движении замыкающих клеток. Каждое устьице имеет две замыкающие специализированные клетки, которые регулируют ширину устьичного отверстия в эпидерме. Под устьичной щелью расположена так называемая 74 Рис. 38. Эпидерма различных растений (вид сверху). А. Хлорофитум (Chlorophytum), представитель однодольных. Б–Г. Представители двудольных (Б – плющ обыкновенный – Hedera helix; В – герань душистая – Pelargonium graveolens: основные эпидермальные клетки – самые крупные, побочные околоустьичные клетки – меньшего размера, они расположены рядом с самыми мелкими замыкающими клетками устьиц, содержащими хлоропласты; Г – шелковица белая – Morus alba). 1 – замыкающая клетка с хлоропластами, 2 – устьичная щель дыхательная полость. Через устьичные отверстия межклетники органов растения сообщаются с атмосферой. Наибольшее число устьиц обычно имеют фотосинтезирующие органы – листья. Однако устьица встречаются и на других надземных органах. Замыкающие клетки устьиц имеют неравномерное утолщение оболочки. Чаще всего к устьичной щели обращена утолщенная оболочка. В состоянии тургора при насыщении замыкающих клеток водой устьичная щель открывается и устанавливается активный газообмен между органом растения и атмосферой. При увядании замыкающих клеток и снижении внутреннего гидростатического давления происходит закрывание устьичной щели. Это приводит к уменьшению транспирации (испарения) и потери воды растением, а также торможению газообмена и связанного с ним процесса фотосинтеза. 75 Рис. 39. Устьица двудольного – А (слива – Prunus), однодольного – Б (рис – Orysa) и голосеменного – В (сосна – Pinus) растений. А1, Б1, Д1. Вид устьиц с поверхности листа (А1 – мицеллы целлюлозы расположены радиально в замыкающих клетках). А2, Б2, В2. Срезы устьиц в плоскости, пересекающей устьичную щель и обе замыкающие клетки. 1 – замыкающая клетка, 2 – побочная клетка, 3 – лигнифицированная часть оболочки, 4 – нелигнифицированная часть оболочки, 5 – углубление в эпидерме над устьицем, 6 – межклетник мезофилла под устьицем, 7 – клетка мезофилла, 8 – клетка эпидермы Расположение замыкающих и побочных клеток, составляющих устьичный аппарат, у различных растений имеет специфические черты, что применяется для их таксономии (рис. 39). В частности, выделяется пять типов устьичных аппаратов: 1) аномоцитный тип (беспорядочный) – устьица не имеют побочных клеток; 2) – анизоцитный тип (неравноклеточный) – устьице окружено тремя побочными клетками, одна из которых меньше, чем две другие; 3) – парацитный (параллельноклеточный) – побочные клетки располагаются параллельно длинной оси устьица; 4) – диацитный (перекрестноклеточный) – устьице окружено двумя побочными клетками, смежные стенки которых расположены поперек длинной оси устьица; 5) – актиноцитный (радиальноклеточный) – несколько побочных клеток окружают устьице, располагаясь радиально относительно замыкающих клеток. 76 Трихомы На поверхности эпидермы образуются разнообразные выросты, называемые трихомами (рис. 40). К ним относятся различные выросты эпидермы – волоски железистые (или секреторные) и неже- Рис. 40. Волоски кроющие и железистые на эпидерме. А. Простые многоклеточные картофеля (Solanum tuberosum). Б. Простые одноклеточные яблони (Malus domestica). В. Ветвистые коровяка (Verbascum thapsus). Г. Многоклеточная звездчатая чешуйка лоха (Elaeagnus angustifolia). Д. Многоклеточные железистые табака (Nicotiana rustica). Е. Этапы развития и скола жгучего эмергенца крапивы (Urtica dioica) лезистые, чешуйки, железки, нектарники. Трихомы могут иметь все органы растений. Трихомы различаются по строению и выполняемым функциям, могут оставаться живыми длительное время или эфемерными, быстро отмирающими. Принято разделять трихомы на кроющие и железистые. 77 Кроющие трихомы могут быть одноклеточными, многоклеточными, ветвистыми и звездчатыми. Железистые волоски имеют одноклеточную или многоклеточную головку, состоящую из секреторных клеток, расположенную на ножке, состоящей из нежелезистых клеток. Железки представляют собой многоклеточные секреторные структуры, расположенные на короткой нежелезистой ножке. Нектарники – это железистые многоклеточные структуры, секретирующие жидкость, содержащую органические вещества и сахара. Трихомы выполняют разнообразные функции, защищают мезофилл от перегрева, механически защищают растения от насекомых, способствуют выведению солей из тканей листа, например у лебеды, и осуществляют химическую защиту. Строение и форма трихом используются для целей систематики растений, а также для микроскопического анализа лекарственного растительного сырья. На эпидерме многих растений образуются многоклеточные выросты – эмергенцы, в образовании которых принимают участие не только поверхностный слой клеток, но и более глубокие их слои. К эмергенцам относятся жгучие «волоски» крапивы, шипы розы, малины, ежевики, плодов зонтичных, дурмана, каштана. Корневые волоски Особый тип трихомов покровной ткани корней – ризодермы (эпиблемы, эпидермы корня) составляют корневые волоски (рис. 41). Это трубчатые живые выросты клеток ризодермы, осуществляющие поглощение воды и разнообразных растворенных веществ, содержащихся в почве. Они образуются в результате разрастания специализированных клеток ризодермы – трихобластов. Корневые волоски сильно вакуолизированы, имеют ядро в пристенном слое цитоплазмы и легко проницаемую оболочку. Корневые волоски достигают в длину 1–2 мм, они многократно увеличивают поглощающую поверхность корня. Продолжительность их жизни составляет 15–20 дней. 78 Развитие корневого волоска из трихобласта сопровождается увеличением числа органелл в цитоплазме этой клетки (рис. 42). Происходит поляризация цитоплазмы. Апикальная часть растущего корневого волоска занята в основном пузырьками аппарата Гольджи, между которыми расположены короткие гранулярные и агранулярные трубочки эндоплазматического ретикулума. Плазмалемма имеет извилистые очертания, что связано с активным процессом обмена веществ между растущей оболочкой и цитоплазмой. Рис. 41. Кончик корня традесканции (Tradescantia). А. Общий вид корневого чехлика, апекса и зоны всасывания, с корневыми волосками. Б. Корневой чехлик и апекс. В. Корневые волоски на разных стадиях развития. Г. Трихобласт с примордием корневого волоска (трихобласты – клетки, образующие корневые волоски, обозначены точками) Рис. 42. Ультраструктура растущего корневого волоска редиса (Raphanus sativus): 1 – оболочка клетки на верхушке корневого волоска, 2 – извилистая плазмалемма, 3 – скопление пузырьков аппарата Гольджи, 4 – гранулярный эндоплазматический ретикулум, 5 – тонопласт вакуоли, 6 – крупная вакуоль, 7 – ядро клетки, 8 – диктиосома, 9 – ядрышко, 10 – наружная поверхность корня, 11 – клетка ризодермы, 12 – клетка первичной коры корня 79 ПЕРИДЕРМА Голосеменные и покрытосеменные растения, обладающие способностью к вторичному радиальному росту, образуют вторичную покровную ткань – перидерму (рис. 43). Перидерма сменяет эпидерму по мере прекращения первичного осевого роста и возникновения вторичного радиального роста проводящего цилиндра стеблей и корней, осуществляемого деятельностью вторичной латеральной меристемы – камбия. Перидерма представляет собой комплексную покровную ткань. Она состоит из наружного покрова пробки – феллемы, сложенной из многочисленных мертвых слоев клеток, оболочки которых пропитаны суберином. Под пробкой находится вторичная боковая образовательная ткань – феллоген, или пробковый камбий. Под ним расположена феллодерма, состоящая из живых паренхимных клеток. Клетки феллогена активно делятся, вычленяя во внешнюю сторону молодые клетки пробки, а во внутреннюю сторону – клетки феллодермы. Феллоген развивается из клеток первичной коры, находящихся под эпидермой и сохраняющих меристематическую активность. Клетки пробки первоначально остаются живыми, имеют таблитчатую форму. По мере созревания их оболочки пропитываются суберином, что придает им непроницаемость для газов и водных растворов. При этом происходит формирование плотного покрова внутренних тканей органа и изоляция внешних живых клеток эпидермы, которые отмирают, оставаясь на поверхности пробки. Побеги при этом изменяют зеленую окраску на бурую. У некоторых растений пробка образует довольно толстые отложения, толщиной в несколько сантиметров, например, у пробкового дуба, бархата амурского. Для нормальной жизнедеятельности живых тканей, лежащих под пробкой, им необходим газообмен с атмосферой. Эту функцию осуществляют чечевички, представляющие собой разрывы в сплошном пробковом покрове, которые заполнены выполняющей тканью. Она состоит из паренхимных тонкостенных слабо опробковевших клеток с широкими межклетниками. Чечевички имеют вид небольших бугорков на поверхности молодых стеблей побегов. С возрастом чечевички разрастаются в направлении длины окружности стебля, например, на стволе березы образуются так называемые «бровки». Перидерма формируется не только на осевых органах, но иногда возникает в кроющих чешуях зимующих почек у голосеменных и двудольных, а также в основаниях черешков листьев осенью. После 80 опадания листа перидерма возникает в месте прикрепления черешка к стеблю, предохраняя последний от высыхания и попадания инфекции. Перидерма возникает также в местах поранения органов растения из клеток раневой меристемы. Рис. 43. Перидерма стебля бузины (Sambucus racemosa). А. Чечевичка. Б. Участок перидермы. 1 – выполняющая ткань, 2 – остатки эпидермы, 3 – пробка (феллема), 4 – пробковый камбий (феллоген), 5 – феллодерма КОРКА В многолетних органах растений перидерма образуется ежегодно путем заложения новых слоев феллогена из живых паренхимных клеток. При этом с внешней стороны от возникающих слоев новой пробки клетки многих тканей, включая и элементы луба, отмирают. В результате этого, происходит накопление многочисленных слоев перидерм и клеток других отмерших тканей, составляющих мощную третичную покровную ткань корку или ритидом (рис. 44). Таким образом, у многолетних растений формируется «годичная» слоистость корки. Стволы деревьев предохраняются коркой от механических 81 повреждений, лесных низовых пожаров, резкой смены температуры, проникновения патогенных организмов. ПРОВОДЯЩИЕ ТКАНИ Обмен веществ составляет основу жизнедеятельности клеток и всего растительного организма. Он осуществляется на молекулярном, внутриклеточном, межклеточном, межтканевом уровнях, а также между различными органами растения. Практически все расположенные рядом живые клетки организма способны осуществлять между собой Рис. 44. Схема строения корки: обмен веществ, обозначаемый 1 – пробка, 2 – паренхима коры, как ближний транспорт ве3 – склереиды, 4 – лубяные волокна, 5 – клетки с друзами кристаллов ществ. Однако в организме существует необходимость быстро доставлять во все ткани и клетки продукты жизнедеятельности от удаленных органов, например от листьев и корней. Эта функция осуществляется единой системой дальнего транспорта веществ, которая расположена во всех органах растения и состоит из проводящих пучков или цилиндров. В их состав входят специализированные проводящие ткани – флоэма и ксилема, как правило, формирующиеся совместно. Это сложные комплексные ткани, состоящие из нескольких типов клеток, специализированных по функции и различающихся морфологически. Ксилема транспортирует в восходящем направлении в надземные органы большие количества воды и растворенных в ней веществ, поглощаемых корнями. ФЛОЭМА Флоэма, или луб, осуществляет дальнюю транспортировку органических веществ от периферических органов – листьев, к осевым органам – стеблям и, далее по ним, к корневой системе. Кроме того, она выполняет функции ближнего транспорта веществ, их накопление, а также придает механическую прочность органам растения. Эти функции осуществляются специализированными типами клеток. 82 В состав флоэмы входят ситовидные элементы – ситовидные клетки, членики ситовидных трубок, клетки-спутники, паренхимные клетки осевой системы, паренхимные и альбуминовые – белковые клетки лучевой системы, а также механические клетки – волокна и склереиды (рис. 45). Основную транспортную функцию во флоэме выполняет особый тип клеток – ситовидные элементы. Главная структурная особенность их состоит в том, что в оболочках они содержат ситовидные участки, которые представляют собой части клеточной оболочки с многочисленными отверстиями; через последние нитевидными тяжами соединяются протопласты двух смежных ситовидных элементов. Каждый тяж в поровом канальце окружен слоем полисахарида – каллозы. Оболочки ситовидных клеток толще, чем оболочки окружающих паренхимных клеток. Они имеют рыхлую структуру, состоят из целлюлозы и пектиновых веществ и не одревесневают. Рис. 45. Типы клеток вторичной флоэмы двудольного растения белой акации (Robinia pseudoacacia). А. Волокно. Б. Членик ситовидной трубки с клетками-спутниками (срез Б2 проходит по плоскости ситовидной пластинки). В. Паренхимные клетки, содержащие кристаллы. Г. Склереида. Д. Тяж осевой флоэмной паренхимы. Е. Клетки луча на тангенциальном (Е1), радиальном (Е2) и поперечном (Е3) срезах. (А1, Б1, В1, Г1, Д1 – продольные срезы; А2, Б2, В2, Г2, Д2 – поперечные срезы). 1 – ситовидная пластинка, 2 – ситовидное поле на боковой стенке членика ситовидной трубки, 3 – клетка-спутник, 4 – кристалл 83 В продолжение всего времени функционирования ситовидного элемента протопласт остается живым. Однако в процессе дифференциации ситовидного элемента происходят его существенные изменения. Ядро деградирует, исчезают диктиосомы и рибосомы. Вакуолярная мембрана – тонопласт растворяется, в результате чего граница между вакуолью и цитоплазмой теряется. Сохраняются пластиды, митохондрии, эндоплазматический ретикулум. В протопластах ситовидных элементов формируются слизевые тельца, состоящие из особого флоэмного белка, обозначаемого как ф-белок. В процессе старения ситовидного элемента каллоза полностью перекрывает отверстия пор ситовидных участков. В результате этого через ситовидные участки прекращается транспортировка веществ. Ситовидные элементы функционируют непродолжительное время, чаще всего в течение одного вегетационного периода. Затем протопласт дезорганизуется и отмирает, а клетка, подвергаясь давлению окружающих тканей, сжимается или растворяется – облитерирует. Общая морфология проводящих клеток флоэмы и структура их ситовидных участков оболочки имеют определенную изменчивость, на основании чего ситовидные элементы подразделяются на ситовидные клетки и членики ситовидных трубок. Ситовидные клетки формируются во флоэме хвойных растений, а ситовидные трубки, состоящие из отдельных клеток члеников ситовидных трубок – во флоэме эволюционно продвинутых покрытосеменных растений (рис. 46). Рис. 46. Типы ситовидных элементов: А – ситовидная клетка, Б – членик ситовидной трубки со сложной ситовидной пластинкой, В – членик ситовидной трубки с простой ситовидной пластинкой 84 Рис. 47. Продольный радиальный срез флоэмы и камбиальной зоны тыквы: 1 – ситовидная трубка, 2 – ситовидная пластинка, 3 – отверстие ситовидной пластинки, 4 – мозолистое тело, 5 – ситовидные поля на боковой стенке ситовидной трубки, 6 – ядро клетки-спутника, 7 – клетки-спутники, 8 – флоэмная паренхима, 9 – ядро камбиальной клетки, 10 – камбиальная зона Ситовидные клетки обычно довольно длинные и узкие. Они имеют ситовидные поля в различных местах продольных стенок, которые не группируются в виде ситовидных пластинок. Членики ситовидных трубок имеют ситовидные поля, сгруппированные в виде ситовидных пластинок на торцевых стенках (рис. 47, 48). Через них сообщаются между собой расположенные друг за другом членики ситовидных трубок. Ситовидные пластинки обеспечивают более эффективный транспорт веществ между члениками ситовидных трубок по сравнению с менее регулярно расположенными на боковых стенках ситовидными полями ситовидных клеток. Транспортировка органических веществ осуществляется протопластами ситовидных элементов и связана со сложными энергозависимыми метаболическими процессами. Однако данные протопласты лишены ядер и не способны самостоятельно осуществлять в полной мере синтез всего комплекса веществ, необходимых для поддержания своей жизнедеятельности. Кроме того, продукты флоэмного транспорта обычно синтезируются на определенном удале85 Рис. 48. Флоэма и камбиальная зона тыквы на поперечном срезе: 1 – ситовидная трубка, 2 – ситовидная пластинка, 3 – клетка-спутник, 4 – отверстие ситовидной пластинки, закупоренное каллозой, 5 – отверстия ситовидной пластинки функционирующей ситовидной трубки, 6 – протопласт ситовидной трубки, 7 – стенка сосуда ксилемы, 8 – камбиальная зона, 9 – паренхима флоэмы нии от ситовидных элементов и должны быть к ним доставлены и загружены, а затем разгружены. Поддержание транспортной функции и жизнедеятельности ситовидных элементов, а также их загрузка и разгрузка органическими веществами осуществляются расположенными рядом с ними клетками-спутниками, передаточными клетками, а также особыми белковыми клетками. Клетка-спутник обычно является дочерней той материнской клетке, от которой происходит и рядом расположенный ситовидный элемент. Клетка-спутник представляет собой паренхимную клетку, тесно связанную плазмодесмами с ситовидным элементом. Она обладает высокой метаболической активностью. Клетка-спутник имеет большое ядро и ядрышки. В плотной гиалоплазме, насыщенной рибосомами, располагаются хорошо развитые пластиды, много крупных митохондрий, эндоплазматический ретикулум, имеются мелкие вакуоли. Вблизи флоэмных окончаний – местах их активной загрузки, например в листе, клетки-спутники имеют внутренние выросты оболочки, смежной с ситовидными элементами. Благодаря этому увеличивается поверхность соприкосновения протопласта клетокспутников с ситовидными элементами и интенсивность обмена 86 Рис. 49. Продольный радиальный срез проводящей зоны вторичной флоэмы сосны обыкновенной (Pinus sylvestris): 1 – ситовидные клетки, 2 – ядра паренхимных клеток, 3 – осевая паренхима, 4 – ситовидные поля, 5 – белковая клетка (клетка Страсбургера), 6 – внутренние лежачие клетки луча, 7 – окончания ситовидных клеток, 8 – радиальный луч Рис. 50. Продольный тангенциальный срез проводящей зоны вторичной флоэмы сосны обыкновенной (Pinus sylvestris): 1 – ситовидные клетки, 2 – ситовидные поля, 3 – ядро клетки сердцевинного луча, 4 – однорядный радиальный луч, 5 – белковая клетка, 6 – многорядный радиальный луч, 7 – кристаллоносная паренхима, 8 – кристаллы оксалата кальция, 9 – осевая паренхима, 10 – радиальный смоляной канал, 11 – эпителиальные клетки, 12 – гранулы крахмала в паренхимной клетке веществ между ними. Такие клетки-спутники с внутренними выростами называют передаточными клетками. У различных покрытосеменных растений клетки-спутники, передаточные и паренхимные клетки флоэмных окончаний имеют определенные особенности структуры, что применяется для таксономии и характеристики их функциональной нагрузки. Около одного ситовидного элемента может находиться несколько клеток-спутников. Они располагаются с одной или нескольких сторон, либо образуют осевые ряды. У голосеменных растений функции клеток-спутников выполняют белковые, альбуминовые клетки, называемые также клетками страсбургера (рис. 49, 50). В отличие от клеток спутников, белковые клетки развиваются из иных инициальных клеток, чем рядом 87 расположенные ситовидные элементы. По цитологическим характеристикам они также напоминают метаболически активные клетки спутники. Белковые клетки дифференцируются из клеток осевой паренхимы или из краевых лучевых клеток – более коротких и высоких по сравнению с обычными клетками лучей. В составе флоэмы функционирует значительное количество живых паренхимных клеток, осуществляющих ближний транспорт и запасание веществ. В них накаливается крахмал, масло, другие органические вещества, а также смолы и танины. Паренхимные клетки образуют осевую и лучевую системы, взаимосвязанные между собой. Осевые паренхимные клетки, обычно прозенхимные, имеют вытянутую форму и происходят из веретеновидных инициалей камбия. Иногда эти клетки делятся поперечными перегородками. В результате этого образуются тяжи осевых паренхимных клеток. Лучи паренхимных клеток формируются из коротких камбиальных лучевых инициалей. Клетки, вытянутые вдоль луча, обозначаются как лежачие, а удлиненные поперек луча – стоячие. В процессе старения флоэмы некоторые паренхимные клетки разрастаются, одревесневают и превращаются в склереиды. Помимо склереид в составе флоэмы присутствуют лубяные волокна. В первичной флоэме волокна образуются на периферии проводящих пучков, а во вторичной – среди проводящих и паренхимных клеток. Волокна имеют вытянутую форму и бывают одноклеточными или многоклеточными – септированными, которые возникают в результате деления одной клетки поперечными перегородками. При созревании клетки волокон отмирают или остаются живыми. Волокна наряду с механической функцией осуществляют запасание веществ. По времени возникновения флоэма подразделяется на первичную и вторичную. Первичная флоэма Первичная флоэма формируется из прокамбия и подразделяется на протофлоэму и метафлоэму. Протофлоэма созревает в процессе интенсивного роста молодых органов. Ситовидные элементы протофлоэмы узкие, относительно слабо дифференцированные, с мало заметными ситовидными пластинками. В протофлоэме часто отсутствуют клетки-спутники. Ее элементы 88 отличаются недолговечностью и по мере роста органа растягиваются и перестают функционировать. Зачатки волокон при этом удлиняются и, созревая, формируют вторичную оболочку. Такие волокна обычно занимают периферическое положение по отношению к протофлоэме. По мере завершения первичного роста органа начинают созревать элементы метафлоэмы. Метафлоэма отличается большим числом ситовидных элементов, которые значительно шире и длиннее, чем в протофлоэме, и имеют более продолжительное время жизни. Ситовидные трубки окружены клетками-спутниками. У однодольных и травянистых двудольных (лютиковых) паренхимные клетки развиваются вокруг сплошных пучков ситовидных трубок с клетками-спутниками. Волокна в метафлоэме обычно не развиваются. Метафлоэма функционирует на протяжении всей жизни однодольных и некоторых двудольных растений, не имеющих вторичного роста. Вторичная флоэма Вторичная флоэма развивается обычно из камбия у голосеменных и двудольных растений, способных формировать эту вторичную латеральную меристему. Он закладывается между первичной флоэмой и первичной ксилемой из остающихся клеток прокамбия, либо из клеток перицикла в корнях. Вертикальная система клеток вторичной флоэмы содержит ситовидные элементы, клетки-спутники или белковые клетки, а также осевую паренхиму, которые формируются удлиненными осевыми камбиальными инициальными клетками. Лучевая система паренхимных клеток развивается из коротких лучевых инициалей камбия. Вторичная флоэма сложена в виде правильных радиальных рядов клеток осевой и лучевой систем, которые продолжаются во вторичной ксилеме. Это связано с тем, что одна и та же камбиальная инициальная клетка, осевая или лучевая, попеременно осуществляет формирование молодых клеток вторичной флоэмы и вторичной ксилемы. В соответствии с характером структуры камбия формируется ярусная или неярусная система клеток осевой и лучевой систем вторичной флоэмы. 89 На поперечном срезе вторичная флоэма иногда имеет слоистость. Это связано с тем, что у некоторых растений клетки вторичного луба, сформированные в начале вегетационного периода, расширяются в тангенциальном направлении, а образующиеся осенью – остаются узкими. Однако такая слоистость не всегда сохраняется из-за отмирания проводящих элементов и преобразования паренхимных клеток в старых частях флоэмы. Обычно клетки вторичной флоэмы осуществляют проводящую функцию в течение одного вегетационного периода. После чего они заменяются новыми слоями молодых клеток. Однако у липы и винограда, например, ситовидные трубки остаются деятельными в течение нескольких лет. У хвойных растений вторичная флоэма устроена относительно однообразно (рис. 49, 50). Она содержит ситовидные клетки, осевую паренхиму, и у многих растений – лубяные волокна. Лучевая система представлена, в основном, однорядными лучами только паренхимных или паренхимных и белковых клеток. Ситовидные клетки имеют ситовидные поля, расположенные, в основном, на радиальных стенках. Они особенно многочисленны у концов клеток. Структура вторичного луба имеет характерные признаки строения у разных хвойных растений, поэтому она может применяться в качестве диагностических признаков в их таксономии. У двудольных растений флоэма имеет более разнообразное строение по сравнению с хвойными растениями (рис. 51, 52). Для флоэмы двудольных характерны клетки-спутники и высоко специа- Рис. 51. Поперечный cрез вторичной флоэмы липы мелколистной (Tilia cordata): 1 – радиальный луч, 2 – поры паренхимной клетки, 3 – лубяные волокна, 4 – ситовидные трубки, 5 – призматический кристалл в паренхимной клетке, 6 – осевая паренхима, 7 – протопласт ситовидной трубки, 8 – клетка спутник, 9 – ситовидная пластинка, 10 – ядро паренхимной клетки, 11 – друза кристаллов в паренхимной клетке 90 лизированные ситовидные трубки. Причем степень дифференциации члеников ситовидных трубок имеет различия у разных таксонов. Мало специализированы ситовидные пластинки, расположенные на боковых стенках члеников ситовидных трубок, у яблоневых. Другие двудольные растения имеют ситовидные пластинки на расширенных поперечных смежных стенках стоящих друг над другом члеников ситовидных трубок (см. рис. 47, 48). Более специализированы ситовидные трубки у травянистых многолетних двудольных по сравнению с древесными растениями. Рис. 52. Продольный радиальный срез вторичной флоэмы липы мелколистной (Tilia cordata): 1 – ситовидная трубка, 2 – кристалл клетки осевой паренхимы, 3 – протопласт ситовидной трубки, 4 – осевая паренхима, 5 – ситовидная пластинка, 6 – лубяные волокна, 7 – ситовидное поле на боковой стенке ситовидной трубки, 8 – клетки спутники, 9 – радиальный луч, 10 – гранулы крахмала В наибольшей мере на структурное разнообразие вторичной флоэмы влияют количество и распределение лубяных волокон. Они могут встречаться в лубе рассеянно, либо в виде полос или отдельных островков. Также разнообразно распределение проводящих элементов флоэмы, ситовидных трубок с клетками-спутниками, которые образуют тангенциальные полосы, чередующиеся с полосами осевой паренхимы у белой акации, или собраны в радиальные ряды у миндаля, либо разбросаны без определенного порядка. В старых частях флоэмы, где обычно отсутствуют нормально функционирующие ситовидные элементы, последние сдавливаются или остаются открытыми и пустотелыми. Клетки лучевой паренхимы продолжают жизнедеятельность, накапливая крахмал, масла, таннины. В связи с увеличением окружности органа в процессе его рос91 та, паренхимные клетки начинают делиться радиальными перегородками. При этом происходит нарастание объема паренхимной ткани в тангенциальном направлении, что обозначается как явление дилатации. Со временем в старых периферических частях луба закладывается феллоген. Он формирует пробку, которая нарушает питание расположенных снаружи от нее слоев клеток, и приводит к их отмиранию. КСИЛЕМА Ксилема (древесина) транспортирует воду и растворенные в ней вещества, поглощаемые из почвы корневыми окончаниями, по корням в основание надземной части растения. Далее по надземной части эти водные растворы перемещаются ксилемой в восходящем направлении к верхушке растения, а также в листья и периферические органы. Весной в растущие органы по ксилеме транспортируются растворы органических веществ, в основном сахаров. Они высвобождаются из крахмала и масла, запасенных в паренхимных тканях корней и стеблей в предыдущий вегетационный период. Наряду с этим, ксилема осуществляет функцию механической ткани, она составляет каркас растения, а также служит местом резервирования воды, крахмала, масла и других запасных веществ. В состав ксилемы входят специализированные клетки, выполняющие определенные функции, включая дальнюю транспортировку веществ, механическое упрочение древесины, накопление веществ и их ближнюю транспортировку. У растений различных таксонов структура ксилемы имеет существенные отличия. Относительно просто устроена ксилема несеменных и голосеменных растений, наибольшего разнообразия и сложности структуры она достигает у покрытосеменных (рис. 53). Все клетки ксилемы разделяются на две основные группы. 1. Трахеальные элементы, включающие трахеиды, членики сосудов, сосудистые трахеиды, волокнистые трахеиды, волокна либ92 риформа. Эти клетки, в основном, удлиннены параллельно продольной оси органов растения. 2. Паренхимные элементы, организованные в две системы – осевую и лучевую. Клетки осевой системы ориентированы в виде слоев или тяжей, расположенных продольно трахеальным элементам. Лучевая система представлена клетками, расположенными в виде радиальных (сердцевинных) лучей. Рис. 53. Типы клеток вторичной ксилемы (древесины) дуба. I–III – широкие членики сосудов, IV–VI – узкие членики сосудов, VII – трахеида, VIII – волокнистая трахеида, IX – волокно либриформа, X – клетки лучевой паренхимы, XI – тяж клеток осевой паренхимы. 1 – лучевые клетки, примыкающие к широкому членику сосуда, 2 – простые перфорации 93 По развитию ксилема подразделяется на первичную ксилему и вторичную. Первичная ксилема формируется из латеральной меристемы прокамбия в процессе развития молодых органов первичного тела растения. Она состоит из двух частей – протоксилемы и метаксилемы. Клетки протоксилемы более ранние в онтогенетическом отношении. Они дифференцируются вместе с активно удлиняющимися тканями в тех частях первичного тела растения, которые еще не полностью закончили рост и дифференциацию. Созревание клеток метаксилемы происходит позже, после того как, в основном, заканчивается удлинение органа. Вторичная ксилема развивается из вторичной латеральной меристемы камбия после завершения первичного роста органов. Трахеальные элементы Трахеальные элементы ксилемы выполняют водопроводящую и опорную функции. Они представляют собой клетки с одревесневшими оболочками, содержащими окаймленные поры (рис. 54). К трахеальным элементам относятся трахеиды и членики сосудов. Трахеиды имеют вид вытянутых клеток с пористыми оболочками. Между трахеидами растворы перемещаются через окаймленные поры. Водопроводящие осевые трахеиды в древесине голосеменных растений в пределах одного годичного слоя отличаются морфологически и функционально. Трахеиды ранней древесины, которые формируются в начале вегетационного периода, – широкие и тонкостенные, они лучше приспособлены к проведению растворов. В поздней древесине трахеиды толстостенные и менее широкие, эти трахеиды в большей мере выполняют механическую функцию. Кроме того, в древесине хвойных формируются трахеиды лучевые, вытянутые перпендикулярно продольной оси органа. Они осуществляют активную транспортировку веществ в направлении вдоль лучей. Трахеиды стали первыми водопроводящими элементами в процессе эволюции наземных растений. Эволюция трахеид у наземных растений происходила в двух противоположных направлениях – в совершенствовании механической и проводящей функций. В результате увеличения толщины стенок и сокращения ширины клеток происходила специализация трахеид на выполнение механических функций. Это привело к формированию волокон, осуществляющих опорную функцию в древесине. 94 В процессе специализации проводящей функции, трахеиды становились короче и шире и, в конечном итоге, превратились в членики сосудов. При созревании члеников сосудов происходило образование сквозных отверстий, через которые растворы стали свободно перемещаться из клетки в клетку. В целом это обеспечило возникновение сосудов – принципиально новых проводящих структур наземных растений более эффективных, чем трахеиды. Сосуды представляют собой сплошные каналы, сложенные из клеток – члеников сосудов, которые расположены в виде осевых рядов. Главная особенность члеников сосудов состоит в том, что в оболочках следующих друг за другом клеток в процессе дифференциации образуется одно или несколько сквозных отверстий – перфораций. Передвижение воды между члениками сосудов осуществляется через эти перфорации. При наличии одного большого отверстия перфорация считается простой, а нескольких отверстий – сложной. Если образуется несколько узких отверстий, расположенных параллельно друг другу на близком расстоянии, то такую перфорацию называют лестничной. Ж А Д Б Е Г В Рис. 54. Типы трахеид и сосудов. А. Трахеида хвойного растения. Б. Сосудистая трахеида покрытосеменного. В. Волокнистая трахеида покрытосеменного. Г. Сосуд голосеменного (эфедра – Ephedra). Д. Сосуд с лестничной перфорацией. Е. Сосуд с простой перфорацией на боковой стенке. Ж. Сосуд с простой перфорацией на поперечной стенке 95 Рис. 55. Направление эволюции члеников сосудов а1–а4: а1 – примитивный тип членика сосуда, характеризуется значительной длиной, сильно скошенными окончаниями с перфорациями лестничного типа и порами, расположенными строго супротивно в направлении, поперечном продольной оси; а4 – короткий и широкий членик сосуда, наиболее приспособленный к транспорту веществ, с простой широкой перфорацией на слабо скошенных окончаниях и порами на боковых стенках, расположенными в очередном порядке. 1 – лестничная перфорация, 2 – супротивное расположение пор, 3 – очередное расположение пор, 4 – простые перфорации В древесине покрытосеменных растений также формируются сосудистые трахеиды, характеризующиеся большим количеством окаймленных пор, но не образующие перфораций, как у члеников сосудов. Членики сосудов эволюционировали от длинных клеток со скошенными концами и перфорациями лестничного типа к коротким широким клеткам с простыми перфорациями (рис. 55). Трахеальные элементы формируют различные типы вторичного утолщения оболочки (рис. 56). Самые ранние элементы первичной ксилемы, возникающие в молодых растущих органах, имеют кольчатые и спиральные утолщения, которые не препятствуют продольному растяжению клеток. Позже витки спирали формируются на близком расстоянии. Лестничное утолщение считается модификацией спирального, которое возникает в результате слияния витков спирали. В результате такого слияния эти утолщения приобретают вид ступеней лестницы. Дальнейшее развитие утолщений вторичной оболочки приводит к образованию сетчатого утолщения, в котором неутолщенные участки оболочки располагаются как ячейки сетки. Наибольшая площадь вторичного утолщения оболочки развивается у трахеальных элементов с пористым типом, при котором оболочка остается тонкой только в области пор. 96 Рис. 56. Схема верхушки молодого побега клещевины (Ricinus) с формирующейся первичной ксилемой – А: 1 – апекс побега, 2 – молодые листья, 3 – основание листа, 4 – пазушная почка (протоксилема в проводящем пучке: 1 – кольчатый сосуд, 2 – спиральный сосуд с расширенными витками, 3 – спиральный сосуд с плотно расположенными витками). Первичная ксилема на поперечном – Б и продольных – В, Г срезах: 5 – протоксилема; 6 – метаксилема; 7 – кольчатое утолщение; 8 – спиральное утолщение; 9 – лестничное утолщение; 10 – сетчатое утолщение; 11 – пористое утолщение 97 Кольчатые и спиральные утолщения вторичной оболочки трахеид относятся к наиболее древнему типу, и отмечены еще у девонских псилофитов (около 300 млн лет тому назад). Затем появляются лестничные и пористые утолщения. Аналогичным образом, в онтогенезе развивающихся органов у высших растений самые первые элементы ксилемы имеют кольчатые и спиральные утолщения, позже появляются сосуды и трахеиды с лестничными и пористыми (точечными) утолщениями вторичной оболочки. Во вторичных оболочках трахеальных элементов имеются простые и окаймленные пары пор. Обычно между соседними трахеидами и сосудами расположено много окаймленных пор, а между волокнами и трахеальными элементами их намного меньше. Между паренхимными клетками и трахеальными элементами образуются полуокаймленные или простые пары пор. Волокна Рис. 57. Волокнистые элементы древесины: 1 – волокнистая трахеида с окаймленными порами, 2 – одноклеточное волокно либриформа с щелевидными порами, 3 – многоклеточное септированное волокно либриформа Снижение водопроводящей функции трахеид и усиление их механической функции у покрытосеменных растений привело к образованию волокнистых трахеид с редкими порами, имеющими окаймление. Последующее эволюционное преобразование у них окаймленных пор в простые щелевидные привело к возникновению волокон либриформа – специализированных механических клеток древесины покрытосеменных растений (рис. 57). У двудольных растений наблюдается значительное разнообразие волокон либриформа по длине, форме окончаний (гладкие, зазубренные, вильчатые и т.д.), Толщине оболочки и наличию или отсутствию спиральных утолщений вторичной оболочки. Волокна могут быть одноклеточными или многоклеточными перегородчатыми – септированными. Наличие септ в волокнах либри98 форма относится к устойчивому морфологическому признаку родов и семейств растений. У растений с легкой древесиной, тополей и ив, оболочки волокон либриформа тонкие; у самшита, кизила, дуба, имеющих твердую древесину, оболочки волокон очень толстые. Паренхимные элементы К паренхимным относятся преимущественно живые клетки древесины, осуществляющие накопление и медленную доставку воды и запасных веществ. Они могут иметь изодиаметрическую или вытянутую форму и содержать крахмал, жиры, таннины, кристаллы. В первичной ксилеме паренхимные клетки слабо дифференцированы, с первичными оболочками. Во вторичной ксилеме паренхимные клетки более специализированы, имеют вторичные утолщения оболочки, а некоторые из них превращаются в склереиды. Во вторичной ксилеме паренхимные элементы образуют две взаимосвязанные системы клеток – лучевую и осевую. Лучевая паренхима слагается из полосок клеток, расположенных радиально в виде лучей, расходящихся от оси органа. Эти лучи также обозначаются терминами радиальные, сердцевинные или древесинные. Лучи формируются клетками лучевых инициалей камбия. Лучи обладают значительной изменчивостью по высоте, ширине, составу различных типов клеток. Простые лучи могут объединяться в сложные. Высота луча определяется на тангенциальных срезах по числу слоев клеток в осевом направлении, а также в абсолютных единицах длины – микрометрах, миллиметрах. У одних растений лучи совсем низкие, однослойные, а для других видов характерны очень высокие лучи, измеряемые сантиметрами, которые сложены несколькими сотнями слоев клеток. Ширина лучей измеряется количеством слоев клеток в тангенциальном направлении и обозначается как рядность. Этот признак обладает значительной устойчивостью у многих видов растений. Так, например, для большинства хвойных и многих родов древесных двудольных характерны только однорядные лучи в сочетании с небольшим количеством двухрядных лучей. Другие роды древесных растений содержат очень широкие лучи, состоящие из нескольких десятков рядов клеток (дуб черешчатый). Существуют также агрегатные, ложно-широкие лучи. Они сложены из сближенных между собой однорядных, двухрядных и трехрядных лучей, которые чаще всего разделены между собой рядами волокон (граб европейский). 99 По мере ежегодного увеличения диаметра камбиальной зоны возникают новые лучи. Это происходит в результате преобразования веретеновидных камбиальных инициалей в лучевые. Существует несколько морфологических типов лучевых клеток: 1) лежачие, вытянутые вдоль луча; 2) стоячие, удлиненные в осевом направлении; 3) квадратные, относящиеся к промежуточному типу. Лучи, состоящие из лежачих и стоячих клеток, обозначаются как гетероцеллюлярные, лучи, включающие только лежачие клетки – гомоцеллюлярные, а сложенные только из стоячих клеток – гомоцеллюлярно-палисадные (рис. 58, 59). В гетероцеллюлярных лучах стоячие клетки могут занимать различное положение, чередоваться слоями с лежачими клетками, либо обрамлять луч сверху и снизу. Агрегатные сердцевинные лучи некоторых двудольных растений состоят из сближенных между собой однорядных, двухрядных или трехрядных лучей, сливающихся или разделенных рядами волокон (рис. 60). Лучи многих растений образуют межклетники в виде лучевых ходов, содержащих смолу, камеди и другие вещества. Функцию вместилищ могут также выполнять отдельные клетки лучей, накапливающие эфирные масла, кристаллы оксалата кальция, воздух и воду. Рис. 58. Радиальные лучи на тангенциальном сечении. А. Гетероцеллюлярный луч. Б. Гомоцеллюлярный однорядный луч. В. Гомоцеллюлярный многорядный луч 100 Рис. 59. Гетероцеллюлярный луч на радиальном разрезе: 1 – стоячие клетки, 2 – лежачие клетки Рис. 60. Агрегатный сердцевинный луч граба (Сarpinus) на тангенциальном разрезе Наряду с лучами паренхимные клетки также группируются в ксилеме в направлении главной оси органа, образуя древесинную или осевую паренхиму. От трахеальных элементов они отличаются наличием округлых простых пор. Осевая паренхима возникает из веретеновидных инициалей камбия. Производная веретеновидной инициали перед формированием в паренхимную клетку камбия проходит ряд клеточных делений. В результате этого из одной камбиальной клетки образуется тяж паренхимных клеток (рис. 61,1). Число клеток в тяжах осевой паренхимы характерно для отдельных видов, родов и семейств. В том случае, когда камбиальная клетка дифференцируется не претерпевая деления, образуется веретеновидная паренхимная клетка (рис. 61, 2). Клетки осевой паренхимы также способны специализироваться, накапливая кристаллы щавелевокислого кальция, эфирных масел, извести и др. При этом они формируются изодиаметрическими и более крупными, чем обычные клетки осевой паренхимы. Такие клетки обозначаются как камерная паренхима древесины (рис. 61, 3). Осевая или древесная паренхима ксилемы распределяется различным образом по отношению к трахеальным элементам. На основании этого различают два основных типа распределения осевой 101 Рис. 61. Типы клеток осевой паренхимы древесины: 1 – тяж осевой паренхимы, 2 – веретеновидная клетка осевой паренхимы, 3 – клетки камерной паренхимы с накоплением друз кристаллов паренхимы: апотрахеальный при котором ее клетки расположены независимо от положения сосудов (рис. 62, I, II), и паратрахеальный, когда клетки осевой паренхимы и сосуды взаимосвязаны (рис. 62, III, IV). Апотрахеальный тип подразделяется, в свою очередь, на диффузную осевую паренхиму, тяжевую и пограничную или маргинальную. Клетки диффузной паренхимы рассеяны одиночно среди волокон по кольцу годичного прироста. Клетки пограничной паренхимы расположены на одном крае кольца годичного прироста (рис. 62, I). Если клетки этого типа находятся в начале годичного прироста, то их обозначают как инициальная осевая паренхима, а в конце его – как терминальная. Тяжевая апотрахеальная осевая паренхима группируется в виде тангенциальных полос (рис. 62, II). Паратрахеальная осевая паренхима подразделяется на следующие типы: скудная вазицентрическая, крыловидная вазицентрическая и сомкнутокрыловидная. Скудная вазицентрическая паренхима образует сплошные обкладки вокруг сосудов. Крыловидная вазицентрическая паренхима располагается вокруг сосудов с образованием тангенциальных выступов (рис. 62, III). У сомкнутокрыловидной вазицентрической паренхимы тангенциальные выросты сливаются в тангенциальные или диагональные тяжи (рис. 62, IV). Клетки паратрахеальной осевой паренхимы древесины метаболически более тесно связаны с трахеальными элементами, чем клетки апотрахеальной паренхимы. В них весной раньше растворяется запасной крахмал в процессе мобилизации углеводов. Освобождающиеся сахара передаются в сосуды для их быстрой транспортировки к почкам. Клетки осевой паратрахеальной паренхимы, тесно взаимосвязанные с сосудами, получили наименование контактных клеток. Они считаются аналогами клетокспутников, обеспечивающих транспортировку сахаров по ситовидным элементам флоэмы. 102 Рис. 62. Схема распределения клеток осевой паренхимы на поперечных срезах древесины. I – пограничная апотрахеальная, II – тяжевая апотрахеальная, III – вазицентрическая крыловидная паратрахеальная, IV – сомкнутокрыловидная паратрахеальная. 1 – сосуд, 2 – сердцевинный луч, 3 – осевая паренхима Паратрахеальный тип распределения осевой паренхимы в эволюционном отношении является более продвинутым, чем апотрахеальный тип, не связанный регулярно с трахеальными элементами ксилемы. Паренхимные клетки древесины сохраняют свою жизнедеятельность продолжительное время. При этом проводящая функция рядом расположенных сосудов постепенно затухает и переходит к молодым годичным слоям, которые формируются из камбия. Полости нефункционирующих сосудов у многих видов растений заполняются выростами из расположенных рядом паренхимных клеток, которые проникают через поры. Эти выросты называются тилами (рис. 63). Ядро и часть цитоплазмы паренхимных клеток Рис. 63. Лучевые клетки, образующие выросты – тилы, которые проникают через поры в полость сосуда у мирта (Myrtaceae). 1 – пора с плазмодесмами; 2 – лучевая клетка; 3 – сосуд; 4 – вторичная оболочка; 5 – первичная оболочка; 6 – тилообразующий слой; 7 – тила 103 переходят в тилу. В результате жизнедеятельности протопласта тилы запасают эргастические вещества, а также образуют вторичные оболочки, которые полностью блокируют проницаемость сосудов. Тилообразование происходит вблизи мест механического повреждения древесины, а также в зоне деятельности дереворазрушающих грибов. Это обеспечивает защиту ксилемы от излишней потери пасоки. С явлением тилообразования связано формирование ядровой древесины. В целом ксилема представляет собой сложно организованный комплекс, состоящий из нескольких типов клеток, обладающих различными физическими свойствами. Регулярная ориентация клеток в ксилеме придает ей свойства типичного анизотропного материала. Анизотропия ксилемы относится к одному из главных понятий древесиноведения. Проводящие пучки Ксилема, как правило, объединена с флоэмой в составе проводящих пучков (рис. 64). Пучки называются коллатеральными (бокобочными) в том случае, когда флоэма в них обращена к внешней Рис. 64. Схематический вид проводящих пучков различного типа (флоэма изображена светлым фоном, ксилема – в виде заштрихованного фона): 1 – коллатеральный (бокобочный), 2 – биколлатеральный, 3 – концентрический центроксилемный (амфикрибральный), 4 – концентрический центрофлоэмный (амфивазальный), 5 – радиальное расположение проводящих тканей (обычно в корнях) стороне органа, а ксилема – к внутренней. Если между этими тканями отсутствует камбий, то такие пучки состоят из первичной флоэмы и первичной ксилемы, и их называют закрытыми коллатеральными. Они закрыты в отношении вторичного радиального роста. Коллатеральные пучки, имеющие камбий между ксилемой и флоэмой, называют открытыми. За счет этой вторичной латеральной меристемы они способны 104 расти в радиальном направлении, увеличивая площадь поперечного сечения вторичной флоэмы и вторичной ксилемы. Существуют биколлатеральные пучки, представляющие собой коллатеральные, в которых с внутренней стороны ксилемы имеется дополнительная флоэма, что характерно для вьюнковых, тыквенных, пасленовых, сложноцветных и др. Эти пучки содержат камбий и относятся к открытому типу. Пучки цилиндрической формы, в которых проводящие ткани расположены сплошным внешним и внутренним слоями, называются концентрическими. В том случае, когда внешний слой занимает флоэма, а внутренний – ксилема, пучок называется центроксилемным (амфикрибральным), а пучок, в котором флоэма расположена по оси, а ксилема во внешнем слое – центрофлоэмным (амфивазальным). В концентрических центрофлоэмных пучках камбий, чаще всего, не образуется. В радиальном типе пучка ксилема и флоэма расположены в виде чередующихся секторов. Такой тип пучка, в основном, формируются в корнях. Обычно ксилема в радиальном пучке расходится лучами от оси, а между ними располагаются флоэма. В том случае, когда ксилема имеет два луча – пучок называют диархным, три – триархным, а много лучей – полиархным. Строение древесины Анатомическое строение древесины принято изучать на срезах трех типов: поперечных, лежащих в плоскостях, перпендикулярных оси стебля; продольных радиальных, расположенных в плоскостях, проходящих через ось стебля; тангенциальных, лежащих в плоскостях, параллельных оси стебля, но не проходящих через нее (рис. 65). Для древесных пород ксилема на этих сечениях имеет ряд общих признаков структуры. На поперечных срезах трахеальные элементы древесины – сосуды и трахеиды разрезаны поперек и имеют изодиаметрическую форму. Четко наблюдаются слои годичных приростов древесины. Лучи разрезаны продольно по узкой стороне. Они следуют в направлении, поперечном границам годичных слоев. 105 Рис. 65. Трехмерная схема вторичной ксилемы cосны веймутовой (Pinus strobus) с обозначением поверхностей поперечного и продольных срезов – тангенциального и радиального На радиальных срезах трахеальные элементы разрезаны продольно и хорошо видна годичная слоистость древесины. Лучи разрезаны продольно по широкой стороне и продолжаются в виде относительно широких полосок паренхимных клеток, расположенных поперек годичных слоев древесины. На тангенциальных срезах трахеальные элементы разрезаны продольно, а лучи – поперек. Лучи выглядят в виде линзовидных включений, длинной стороной расположенных продольно оси стебля. 106 Древесина хвойных растений Основной объем древесины хвойных растений сложен осевыми водопроводящими трахеидами. В состав лучей входят лучевые трахеиды. Паренхимные клетки организованы, главным образом, в виде лучей. Кроме того, в древесине имеется сопровождающая паренхима смоляных каналов, а также клетки эпителия, окружающие полость смоляных каналов. Клетки осевой паренхимы встречаются лишь у некоторых хвойных, к которым относятся секвойя и таксодиум. У хвойных пород на полях перекреста в местах контакта паренхимных клеток лучей и осевых трахеид встречается пять типов пор (рис. 66). Рис. 66. Поры на полях перекреста в контакте лучей и осевых трахеид хвойных растений: 1 – купрессоидные поры, 2 – таксодиоидные поры, 3 – пицеоидные поры, 4 – пиноидные поры, 5 – оконцевые поры Купрессоидные окаймленные поры характерны для древесины кипарисовых. У этих пор внутреннее щелевидное отверстие – апертура поры имеет длину меньшую, чем диаметр наружной апертуры. Таксодиоидные окаймленные поры характерны для древесины тисса. Данные поры имеют внутреннюю апертуру, по длине равную диаметру внешней апертуры, а окаймление сохраняется только по бокам поры. Пицеоидные окаймленные поры характерны для древесины ели. У этого типа окаймленных пор внутреннее отверстие еще более увеличенное, чем у таксодиоидных пор. Пиноидные поры относятся к простым порам и не имеют окаймления. Они встречаются небольшими группами на одном поле перекреста, например у древесины сосны желтой. Оконцевые простые широкие поры имеют большую площадь и близкие к четырехугольным очертания. Они расположены пооди107 ночно на одном поле перекреста. Такие поры характерны для сосен и некоторых других родов хвойных, например, подокарповых. Типы пор на полях перекреста у хвойных имеют очень большое диагностическое значение, особенно при определении ископаемых древесин этой группы. Д р е в е с и н а с о с н ы о б ы к н о в е н н о й (Pinus sylvestris) У сосны обыкновенной, как и у большинства хвойных растений, древесина устроена относительно просто. Основной объем ее сложен из осевых трахеид ранней и поздней древесины, выполняющих водопроводящую и механическую функции. Это длинные клетки без протопластов, в оболочках которых имеется много окаймленных пор. На краях лучей находятся лучевые трахеиды, удлиненные поперек оси стебля. В древесине содержатся живые паренхимные клетки, которые входят в состав лучей. Кроме того, живые клетки входят в состав сопровождающей паренхимы смоляных каналов. Полости смоляных каналов окружены клетками эпителия. Смоляные каналы двух типов – осевые и радиальные. На радиальном сечении осевые трахеиды, разрезанные продольно, имеют вид длинных клеток (рис. 67). Раннюю древесину образуют широкие трахеиды с крупными окаймленными порами, Рис. 67. Продольный радиальный срез древесины сосны обыкновенной (Pinus sylvestris): 1 – луч, 2 – осевые трахеиды поздней древесины, 3 – осевые трахеиды ранней древесины, 4, 11 – отверстие (апертура) больших окаймленных пор, 5 – оконцевая (окновидная) пора, 6 – лучевая паренхима, 7 – лучевые трахеиды, 8 – малая окаймленная пора, 9 – торус, 10 – наружный край утолщения вторичной оболочки большой окаймленной поры, 12 – окончания осевых трахеид 108 позднюю – узкие с мелкими окаймленными порами в виде прозрачных колец. В поздней древесине содержатся осевые смоляные каналы в виде длинных узких полостей, ориентированных продольно осевым трахеидам, которые ограничены эпителиальными клетками и клетками сопровождающей паренхимы. Паренхимные клетки также находятся в средней части лучей. На полях перекреста в древесине сосны расположены оконцевые поры. Это широкие простые прямоугольные поры, через которые паренхимные клетки луча сообщаются с осевыми трахеидами. Паренхимные клетки по краям луча ограничены лучевыми трахеидами, имеющими мелкие окаймленные поры. Внутренний край их клеточной оболочки сильно извилистый, зубчатый, что характерно для древесины сосны. На тангенциальном сечении трахеиды разрезаны продольно (рис. 68). Их окаймленные поры расположены в профиль и выглядят в виде чередующихся утолщений в оболочке трахеид. Вертикальные смоляные каналы имеют такой же вид, как и на радиальном сечении. Рис. 68. Продольный тангенциальный срез древесины сосны обыкновенной (Pinus sylvestris): 1 – лучевые трахеиды, 2 – эпителиальные клетки, 3 – полость лучевого смоляного канала, 4 – сопровождающая паренхима многорядного луча, 5 – торус, 6 – оконцевая пора, 7 – лучевая паренхима узкого луча, 8 – окончание осевой трахеиды, 9 – оболочка паренхимной клетки луча, 10 – окаймленная пора 109 Лучи разрезаны поперек, в большинстве – узкие однорядные. В широких лучах расположены лучевые смоляные каналы. Полость такого канала в виде небольшого круглого отверстия в луче ограничена эпителиальными клетками, которые окружены клетками сопровождающей паренхимы. Оконцевые поры на полях перекреста расположены в профиль. Они обнаруживаются по участкам тонкой вторичной оболочки водопроводящих трахеид. Годичной слоистости древесины на тангенциальном срезе не наблюдается. На поперечном сечении осевые трахеиды разрезаны поперек и выглядят в виде прямоугольных клеток (рис. 69). В ранней древесине трахеиды более широкие, в поздней – узкие. Окаймленные поры расположены в профиль. Хорошо видна годичная слоистость древесины. Вертикальные смоляные каналы находятся Рис. 69. Поперечный срез древесины сосны обыкновенной (Pinus sylvestris): 1 – осевые трахеиды ранней древесины, 2 – окаймленные поры, 3 – луч, 4 – оконцевая пора, 5 – торус окаймленной поры, 6 – граница годичного прироста древесины, 7 – капли смолы, 8 – осевые трахеиды поздней древесины, 9 – полуокаймленная пора, 10 – сопровождающая паренхима, 11 – полость осевого смоляного канала, 12 – эпителиальные клетки, 13 – лучевая паренхимная клетка, 14 – лучевая трахеида 110 в поздней древесине. Они разрезаны поперек, в виде округлых отверстий, выстланных эпителиальными клетками, которые окружены клетками сопровождающей паренхимы. Лучи разрезаны продольно по узкой стороне. Они имеют вид рядов в один слой клеток, расположенных поперек годичных слоев древесины. Оконцевые поры ориентированы в профиль и обнаруживаются по значительному сужению вторичной оболочки осевых трахеид. В осевой части ствола у сосны образуется окрашенная ядровая древесина, которая окружена светлой древесиной заболони. Древесина л и с т в е н н и ц ы (Larix) Лиственница имеет твердую прочную и стойкую к разложению древесину, которая широко используется для кораблестроения, в подземных работах, а также в строительстве. В древесине лиственницы содержатся следующие разновидности клеток: осевые водопроводящие трахеиды, выполняющие проводящую и механическую функции; лучевые трахеиды, расположенные на краях лучей (мертвые тонкостенные клетки с мелкими окаймленными порами); паренхимные живые клетки, входящие в состав лучей и осевых смоляных каналов; клетки сопровождающей паренхимы, окружающие эпителиальные клетки смоляных ходов; эпителиальные секреторные клетки, окружающие полости смоляных вместилищ. Основные диагностические признаки древесины лиственницы. Макроскопический признак – наличие в осевой части ствола дерева темного ядра, вокруг которого находится светлая древесина, называемая заболонью. Микроскопические признаки: наличие пицеоидных пор на полях перекреста; резкий переход от ранней к поздней древесине; присутствие тангенциальных групп осевых смоляных каналов в поздней древесине; двойные ряды крупных окаймленных пор в осевых трахеидах ранней древесины; наличие крассул или балок санио – утолщений срединной пластинки и первичной оболочки, расположенных в виде штрихов поперек длинной оси водопроводящих трахеид. Радиальный срез древесины лиственницы характеризуется тем, что на нем наблюдается четкий переход от широких осевых трахеид ранней древесины к весьма узким и толстостенным трахеидам поздней древесины (рис. 70). Трахеиды ранней древесины имеют крупные окаймленные поры, которые расположены на радиальных стенках преимущественно в два ряда, параллельных продольной оси клеток. На этих же стенках находятся крассулы – утолщения срединной пластинки и первичной оболочки. Они имеют вид прозрачных штрихов, ориентированных поперек продольной оси трахе111 ид. Крассулы лучше заметны при закрытой диафрагме конденсора микроскопа в параллельных лучах света. Трахеиды поздней древесины имеют толстую оболочку, узкую полость и мелкие окаймленные поры. Поэтому их транспортная Рис. 70. Радиальный срез древесины лиственницы (Larix): 1 – осевая трахеида поздней древесины, 2 – осевая трахеида ранней древесины, 3 – крупная окаймленная пора в стенке осевой трахеиды, 4 – крассула в стенке осевой трахеиды, 5 – радиальная трахеида в составе луча, 6 – мелкая окаймленная пора в оболочке радиальной трахеиды, 7 – паренхимная клетка луча с пицеоидными окаймленными порами, соединяющими ее с осевыми трахеидами, 8 – мелкая окаймленная пора в оболочке осевой трахеиды поздней древесины функция заторможена, они в основном обеспечивают механическую прочность древесины. Паренхимные клетки лучей на полях перекреста сообщаются с осевыми трахеидами ранней древесины при помощи многочисленных пицеоидных пор. Лучевые трахеиды удлинены вдоль луча. Они имеют относительно ровную толщину оболочки, в которой встречаются мелкие окаймленные поры. На тангенциальном срезе древесины лиственницы в ранней части годичного слоя в осевых трахеидах крупные окаймленные поры расположены в профиль (рис. 71). Они выглядят в микроскопе в виде прозрачных расширяющихся утолщений оболочки, расположенных как 112 Рис. 71. Тангенциальный срез древесины лиственницы (Larix): 1 – полость осевой трахеиды, 2 – крупные окаймленные поры в стенке осевой трахеиды, 3 – радиальная трахеида в узком луче, 4 – паренхимная клетка узкого луча, 5 – паренхима широкого луча, 6 – секреторная клетка радиального смоляного вместилища, 7 – полость радиального смоляного вместилища бусины, нанизанные на нити. Особенно много таких пор в местах окончания, выклинивания трахеид, что способствует осевому транспорту ксилемных потоков веществ. Лучи преимущественно узкие, имеют вид однорядных полосок клеток, расположенных параллельно продольной оси водопроводящих трахеид. На верхнем и нижнем краях лучей находятся лучевые трахеиды треугольного сечения. Паренхимные клетки лучей – с относительно утолщенными оболочками. Многорядные лучи содержат по одному радиальному смоляному каналу, окруженному эпителиальными клетками. Полость такого канала выглядит в виде овального отверстия в средней части луча. На поперечном срезе древесины лиственницы осевые водопроводящие трахеиды выглядят в виде многогранных изодиаметрических клеток (рис. 72). Они расположены правильными радиальными 113 Рис. 72. Поперечный срез древесины лиственницы (Larix): 1 – полость осевой трахеиды ранней древесины, 2 – окаймленные поры в стенке осевой трахеиды ранней древесины, 3 – граница годичного слоя, 4 – осевая трахеида поздней древесины, 5 – секреторная клетка осевого смоляного канала, 6 – полость смоляного осевого канала, 7 – луч рядами, каждый из которых происходит от одной инициальной клетки камбия. На данном срезе хорошо заметен резкий переход от ранней к поздней части годичного слоя. В тонкостенных широких трахеидах ранней древесины на радиальных частях оболочек видны в профиль крупные окаймленные поры. Очень часто они располагаются парами, поскольку эти поры образуют в трахеидах двойные продольные ряды. Клетки лучей на данном сечении имеют удлинение в радиальном направлении. Осевые смоляные каналы обычно располагаются в поздней части годичного слоя. Их округлая полость окружена эпителиальными клетками, вырабатывающими живицу и секретирующими ее в канал. На границе годичного слоя трахеиды поздней древесины наиболее уплощены в радиальном направлении. Трахеиды ранней древесины следующего года – более тонкостенные и вытянуты в радиальном направлении, что связано с их способностью к активному росту в начале вегетационного периода. 114 Древесина покрытосеменных растений Д р е в е с и н а л и п ы м е л к о л и с т н о й (Tilia cordata) Древесина липы состоит из сосудов (трахей) и трахеид, выполняющих функцию транспортировки веществ, древесинных волокон, или волокон либриформа, обеспечивающих механическую прочность древесины, тяжевой и лучевой древесинной паренхимы, осуществляющей накопление и ближний транспорт веществ. На поперечных срезах древесины (рис. 73) границы между кольцами годичного прироста выражены хорошо, так как периферическая часть поздней древесины составлена трахеальными клетками, сжатыми в радиальном направлении. Эти клетки более толстостенные по сравнению с элементами ранней древесины, отложенными камбием в начале вегетационного периода. Рис. 73. Поперечный срез древесины липы (Tilia cordata) на границе двух следующих друг за другом колец годичных приростов: 1 – луч, 2 – трахеида, 3 – граница годичного прироста древесины, 4 – волокна либриформа, 5 – осевая паренхима, 6 – сосуды, 7 – часть спирального утолщения стенки сосуда 8 – окаймленная пора, 9 – простые поры: (1,2 – структуры поздней древесины, 4–8 – структуры ранней древесины) 115 Липа относится к числу древесных пород с рассеяннососудистой древесиной, так как сосуды – наиболее широкопросветные элементы древесины, расположены диффузно по кольцу годичного прироста (рис. 73). Число сосудов и размеры их поперечного сечения постепенно уменьшаются в направлении от ранней к поздней части годичного слоя. На поперечных срезах сосуды имеют овальные, округлые или округло-многоугольные очертания. Сосуды имеют многочисленные мелкие окаймленные поры, с овальными окаймлениями и узкими щелевидными отверстиями, а также спиральные утолщения, возникающие из вторичной оболочки. Такие сосуды называют спирально-пористыми. Их структурные особенности хорошо видны на продольных срезах древесины. Членики сосудов сообщаются между собой посредством простых (одиночных) перфораций, отверстий, имеющих овальные или округлые очертания и расположенных обычно наклонно к продольной оси сосуда. Трахеиды встречаются, главным образом, в поздней древесине. По сравнению с сосудами они имеют более узкий свободный просвет. На поперечных срезах трахеиды имеют многоугольную форму. На продольных срезах они представляют собой длинные прозенхимные клетки со спиральными, часто перекрещивающимися утолщениями внутренних слоев клеточных стенок. Либриформ, или древесинные волокна, это длинные прозенхимные многогранные клетки с гладкими оболочками. В поздней древесине волокна часто имеют таблитчатые очертания поперечного сечения. Как правило, они представляют собой наиболее толстостенные элементы древесины. Однако, по сравнению с другими породами, липа имеет довольно тонкостенные волокна, поэтому ее древесина мягкая и характеризуется малой объемной массой. Стенки древесинных волокон снабжены мелкими щелевидными порами без окаймления. Они отчетливо заметны на продольных срезах. В смежных стенках соседних клеток длинные оси щелевидных каналов каждой пары пор по-разному ориентированы по отношению к продольным осям клеток. При фокусировке с помощью микрометрической рукоятки микроскопа можно видеть, что щелевидные отверстия поровых каналов перекрещиваются, и поры в стенках либриформа выглядят крестовидными. Клетки тяжевой, осевой древесинной паренхимы на поперечных срезах расположены диффузно или собраны в тангенциальные цепочки из двух-трех клеток. На продольных срезах (рис. 74) видно, что каждый тяж состоит из нескольких вытянутых в длину клеток с 116 Рис. 74. Продольный радиальный срез древесины липы (Tilia cordata): 1 – волокно либриформа, 2 – окаймленная пора, 3 – трахеида, 4 – щелевидная пора, 5 – спиральные утолщения вторичной оболочки трахеиды, 6 – простая перфорация сосуда, 7 – крестовидные поры, 8 – спирально-пористый сосуд, 9 – луч, 10 – простые поры, 11 – осевая паренхима пористыми стенками. Как и в лубяной паренхиме, конечные клетки тяжей древесинной паренхимы клиновидно заострены. В тяжевой паренхиме откладываются запасной крахмал или масло. Клетки лучевой паренхимы слегка вытянуты в радиальном направлении. Структурные особенности древесинных лучей лучше всего выражены на радиальных срезах (рис. 74). У липы лучи гомогенные, так как состоят из морфологически одинаковых клеток. Лучевая паренхима выполняет функцию запасания веществ и участвует в их перемещении в радиальном направлении. Диагностические признаки древесины липы: рассеянно-сосудистая древесина; сосуды имеют простые перфорации; сосуды и трахеиды – со спиральными утолщениями оболочки; лучи узкие, двухтрехрядные, высотой до 30 слоев клеток. 117 Д р е в е с и н а д у б а ч е р е ш ч а т о г о (Quercus robur) В состав древесины дуба входят следующие типы клеток: сосуды, сосудистые трахеиды, трахеиды, волокнистые трахеиды, волокна либриформа, паренхима лучей, осевая паренхима. Сосуды представляют собой трубчатые элементы, которые формируются крупными широкими клетками – члениками сосудов, расположенными в виде осевых рядов. В процессе дифференциации члеников сосудов их смежные оболочки растворяются с образованием отверстий – перфораций. Крупные сосуды видны невооруженным глазом. Они находятся в ранней части годичных слоев прироста древесины. Мелкие сосуды расположены в средней и поздней частях годичных слоев группами, которые ориентированы параллельно лучам. Сосудистые трахеиды отличаются от настоящих сосудов тем, что между стоящими друг над другом клетками сохраняются их оболочки с порами. Транспортировка веществ по ним несколько замедлена в сравнении с настоящими сосудами. Трахеиды обычно сопровождают мелкие и крупные сосуды. Они представляют собой замкнутые удлиненные клетки, снабженные большим количеством окаймленных пор. Волокнистыми трахеидами называются такие трахеиды, которые имеют немного пор с окаймлением. Волокна либриформа представляют собой клетки механической ткани. Они узкие, длинные, толстостенные, с мелкими щелевидными порами, без окаймления. Волокна образуют массивные тангенциальные прослойки, которые придают древесине дуба высокую прочность. Лучевая паренхима организована в виде узких и широких лучей. Узкие однорядные лучи встречаются довольно часто. Широкие лучи имеют в ширину до 30 рядов клеток и встречаются реже, чем узкие лучи. Осевая древесинная паренхима сложена в виде тангенциальных прослоек, параллельных продольной оси ствола. Ее клетки контактируют с лучами, образуя единую систему живых паренхимных клеток древесины. Они выполняют запасающую функцию и осуществляют ближнюю осевую и радиальную транспортировку веществ. 118 Дуб имеет кольцесосудистую древесину. Она характеризуется тем, что в ранней части кольца годичного прироста древесины расположены крупные сосуды (рис. 75). В старой древесине около оси ствола проводящая функция сосудов затормаживается. В полости таких сосудов через их поры прорастают паренхимные клетки, расположенные рядом. Выросты этих клеток – тилы заполняют сосуды. По мере вторжения тилы в полость сосуда замыкающая пленка поры разрушается ферментами, в результате чего в полости сосуда тила раздувается в виде шара. Ядро и часть цитоплазмы паренхимной клетки обычно мигрируют в тилу. Тилы способны запасать эргастические вещества, откладывать вторичные оболочки и даже дифференцироваться в склереиды. Тилы закупоривают полость сосудов и снижают проницаемость древесины. С образованием тил связано возникновение ядровой древесины. На поперечном срезе четко обнаруживаются годичные слои прироста (рис. 75). Крупные сосуды, расположенные в ранней древесине, имеют овальное сечение. В их полости расположены тилы. Крупные сосуды окружают трахеиды. Узкие однорядные лучи, не прерываясь, огибают крупные сосуды. Широкие лучи на поперечном срезе древесины имеют относительно толстостенные вытянутые паренхимные клетки. Рис. 75. Поперечный срез древесины дуба черешчатого (Quercus robur): 1 – крупный сосуд в ранней древесине, 2 – тилы в крупном сосуде, 3 – узкий луч, огибающий сосуд, 4 – трахеида, 5 – поздняя древесина, 6 – мелкий сосуд в поздней древесине, 7 – граница слоя годичного прироста древесины, 8 – широкий луч, 9 – волокна либриформа 119 Рис. 76. Радиальный срез древесины дуба черешчатого (Quercus robur): 1 – сосудистая трахеида, 2 – перегородка с простыми порами между сосудистыми трахеидами, расположенными друг над другом, 4 – клетка осевой паренхимы, 5 – клетка узкого луча, 6 – волокно либриформа Волокна либриформа образуют довольно многочисленные скопления между сосудами и лучами в виде мелких многогранных изодиаметрических клеток с узкой полостью и толстыми оболочками. Мелкие сосуды видны в поздней древесине вблизи границы годичного слоя. Трахеальные элементы на радиальном и тангенциальном продольных срезах древесины дуба выглядят как относительно широкие длинные полости (рис. 76, 77). Сосудистые трахеиды, так же как и сосуды, окружены обычными узкими трахеидами с окаймленными порами. Трахеиды имеют вид длинных клеток с клиновидными окончаниями. В отличие от предыдущих, волокна либриформа более толстостенные и не имеют окаймленных пор. На радиальном срезе видны лежачие клетки луча, длинной осью расположенные поперек трахеальных элементов. На тангенциальном срезе лучи разрезаны поперек. Однорядные лучи имеют вид рядов, изодиаметрических относительно толстостен120 Рис. 77. Тангенциальный срез древесины дуба черешчатого (Quercus robur): 1 – полость мелкого сосуда, 2 – узкий луч, 3 – трахеиды с окаймленными порами, 4 – волокна либриформа, 5 – клетка осевой паренхимы, 6 – широкий луч ных клеток, расположенных продольно трахеальным элементам. Многорядный луч представлен массивом изодиаметрических клеток, также продолжающимся вдоль трахеальных элементов. Древесина дуба имеет следующие диагностические признаки. Кольцесосудистая порода. Наличие крупных сосудов. Наличие широких и узких лучей. МЕХАНИЧЕСКИЕ ТКАНИ Молодые органы растений, сложены живыми клетками с тонкими оболочками. Они сохраняют свою форму, ориентацию в пространстве и способность к росту в различных направлениях, благодаря наличию в клетках тургора – внутреннего гидростатического давления. Возникновение тургора связано со способностью клеток, поглощать и удерживать воду путем создания достаточного осмотического потенциала. Величина осмотического потенциала предопределяется концентрацией растворенных веществ, которую в некотором диапазоне клетки способны регулировать. При значительном недостатке воды тургорное давление 121 снижается до нуля, и клетки теряют свою упругость. Это приводит к увяданию тканей и органов растения. Такое состояние клеток отрицательно сказывается на многих процессах жизнедеятельности, в том числе и на их росте. Механические ткани поддерживают устойчивую форму и ориентацию в пространстве органов растений, их различных тканей, а также полостей – межклетников, заполненных воздухом или продуктами выделений, на фоне периодического увядания. Механические ткани выполняют опорную функцию и придают устойчивость различным органам растения к статическим, постоянным, и динамическим, переменным, механическим нагрузкам. Механические ткани подразделяются на колленхиму и склеренхиму. КОЛЛЕНХИМА Колленхима развивается в молодых и растущих в длину органах растений – под эпидермой стеблей, в черешках листьев. Это самая первая по времени возникновения механическая ткань. Ее клетки имеют живой протопласт, способны выполнять функции обмена веществ, включая фотосинтез, поскольку часто содержат нормально развитые хлоропласты. Оболочки клеток колленхимы имеют неравномерную толщину, как правило, состоят из целлюлозы и богаты пектинами. Они эластичные, что обеспечивает нормальный рост молодых органов в различных направлениях. Клетки колленхимы обычно удлинены параллельно продольной оси органа. Они способны выполнять механическую функцию в состоянии тургора. В зависимости от характера утолщения клеточной оболочки, колленхима подразделяется на уголковую, пластинчатую и рыхлую. Клетки уголковой колленхимы имеют значительное утолщение оболочки только в углах, в местах соединения нескольких клеток (рис. 78). На поперечном срезе такие утолщения выглядят в виде треугольников или ромбов. Пластинчатая колленхима состоит из клеток, имеющих утолщенные тангенциальные стенки, параллельные наружной поверхности органа, а их радиальные стенки значительно менее толстые (рис. 79,а). 122 Рыхлая колленхима образована клетками, между которыми имеются пустотелые межклетники. Характерно, что наиболее утолщенная оболочка формируется вблизи межклеточных пространств (рис. 79,б). СКЛЕРЕНХИМА Механическая ткань, сложенная из клеток, имеющих равномерное утолщение целлюлозных или одревесневших оболочек, называется склеренхимой. Оболочки склеренхимных клеток чаще всего возникают в результате вторичного утолщения. Рис. 78. Уголковая колленхима на поперечном срезе черешка листа бегонии (Begonia): 1 – клетка эпидермы, 2 – клетка колленхимы, 3 – хлоропласт в клетке колленхимы, 4 – утолщение оболочки смежных клеток колленхимы, 5 – клетка паренхимы Рис. 79. Поперечный срез колленхимы: А – пластинчатая, Б – рыхлая. 1 – срединная пластинка клеточной оболочки, 2 – утолщенная клеточная оболочка, 3 – тонкий участок клеточной оболочки, 4 – межклетники, 5 – вакуоль, 6 – хлоропласты В зависимости от формы клеток различают два типа склеренхимы – волокна и склереиды. Волокна представляют собой значительно вытянутые узкие клетки. Склереиды имеют разнообразные очертания и относительно небольшие различия по величине диаметра в трех направлениях. 123 Волокна Волокна располагаются в различных частях тела растения в виде пучков или сплошными цилиндрическими слоями в коре, флоэме, а также группами или рассеянно – во флоэме и ксилеме. В связи с этим волокна разделяются на две группы: древесинные, или ксилемные, и не связанные с древесиной – интерксилярные. Волокна, не связанные с древесиной, подразделяются на следующие типы: 1) лубяные волокна расположены в первичной или вторичной флоэме; 2) коровые волокна, развивающиеся в коре; 3) периваскулярные волокна, формирующиеся по периферии пучков проводящих тканей, но возникающие независимо от ксилемы и флоэмы. Считается, что периваскулярные волокна возникают из перицикла, расположенного между корой и проводящими тканями. Интерксилярные волокна развиваются различным путем. В коре они закладываются из клеток меристемы основной паренхимы, которые перестают делиться поперечными перегородками и начинают удлиняться. Волокна, относящиеся к первичной флоэме или окружающие проводящие пучки и состоящие из первичных проводящих элементов, возникают из прокамбия. Во вторичной флоэме лубяные волокна формируются из веретеновидных инициальных клеток камбия, а также в результате преобразования клеток паренхимы луба. Интерксилярные волокна могут состоять только из целлюлозы, например у льна (рис. 80), или быть значительно одревесневшими, что характерно для однодольных растений. Первичные волокна луба, возникающие из прокамбия, достигают значительно большей длины, чем вторичные, формируемые камбием. Например, у конопли, первичные волокна имеют длину около 12,7 мм, а вторичные – 2,2 мм. При этом длинные, первичные, волокна формируются многоядерными клетками, а вторичные – одноядерными. Наибольшей длины волокна достигают крапивы рами (Boehmeria nivea). Лубяные волокна представляют собой важное сырье для изготовления технического волокна, играющего большую роль в экономике. Технические волокна получают, подвергая растения процессу мочки, в котором участвуют 124 Рис. 80. Поперечный разрез стебля льна обыкновенного (Linum usitatissimum) с первичными флоэмными волокнами: 1 – эпидермис, 2 – кора, 3 – флоэмные волокна, 4 – флоэма, 5 – камбий, 6 – вторичная ксилема бактерии и грибы, разрушающие пектиновые вещества межклеточного материала. В результате этого пучки волокон освобождаются от окружающих клеток, становятся эластичными и пригодными для изготовления пряжи, используемой при получении тканей, веревок, канатов и т.п. Лен (Linum usitatissimum) наиболее известен с давних пор как текстильное растение, имеющее лубяные волокна. Возделывался еще в Древнем Египте, Китае и других странах, более 5000 лет назад. Кроме льна широко применяются лубяные волокна конопли (Cannabis sativa), джута (Corchorus capsilaris), рами (Boehmeria nivea), кенафа (Hibiscus cannabinus) и других растений. Для лубяных волокон характерно наличие слоистости вторичной оболочки. 125 Волокна древесины характеризуются морфологическим разнообразием, но имеют общее происхождение – из камбия (см. рис. 57). Типичные волокна древесины, сходные по структуре с флоэмными, принято обозначать волокнами либриформа. Кроме того, в древесине свойства специализированных механических клеток приобретают волокнистые трахеиды, представляющие собой трахеиды, лишенные возможности осуществления активной транспортировки веществ из-за наличия очень редких окаймленных пор. Волокна древесины и луба иногда имеют поперечные перегородки, которые возникают в результате деления первичной клетки волокна, возникающей из веретеновидной камбиальной инициали. Склереиды Склереиды представляют собой клетки механической ткани, имеющие значительное утолщение вторичной одревесневшей оболочки, пронизанной многочисленными, обычно простыми, порами. Склереиды широко распространены в растениях и встречаются во всех их органах. Они встречаются в виде скоплений среди других клеток. Одиночные склереиды обозначаются как идиобласты. Склереиды входят в состав различных структур стеблей – эпидермы, первичной коры, проводящих тканей, сердцевины, мезофилла листа, покровных и основных тканей плодов и семян. Склереиды формируются из клеток апикальных и латеральных меристем или в результате дифференциации уже зрелых паренхимных клеток. Форма склереид весьма разнообразна и по этому признаку они разделяются на несколько типов. Каменистые клетки, относительно изодиаметрические, обозначаются как брахисклереиды; палочковидные удлиненные клетки – макросклереиды; клетки, имеющие утолщения на двух концах наподобие трубчатой кости – остеосклереиды; клетки с лопастями, отходящими от центральной части наподобие звезды – астросклереиды; клетки, подобные волоскам растений – трихосклереиды; клетки длинные, по виду напоминающие волокна – нитевидные склереиды (рис. 81). 126 Рис. 81. Склереиды. А, Б – каменистые клетки из мякоти плода груши – Pyrus; В, Г – склереиды из первичной коры стебля воскового плюща – Hоуа (В – вид на срезе, Г – вид с поверхности); Д, Е – склереиды эндокарпия плода яблони – Malus; Ж – колончатая склереида с разветвленными концами из палисадного мезофилла – Hakea; 3, И – склереиды из черешка Camellia; К – астросклереида из первичной коры стебля Trochodendron; Л – слой эпидермальных склереид в луковичной чешуе чеснока – Allium sativum; M, Н – нитевидные склереиды из мезофилла листа маслины – Olea; О–Р – склереиды из субэпидермального слоя семенной кожуры фасоли – Phaseolus, имеющие форму песочных часов (О, П – вид сбоку, Р – вид сверху); С, Т – эпидермальные склереиды – макросклереиды – с выемчатыми утолщениями оболочки из семенной кожуры фасоли – Phaseolus (C – вид сбоку, Т – вид сверху). 1 – ветвистая пора, 2 – простая пора, 3 – полость клетки 127 ОСНОВНЫЕ ТКАНИ В органах растения основная ткань занимает наибольший объем и представлена, главным образом, паренхимой. Она образует непрерывную ткань и составляет связующее звено между другими тканями. Ее клетки специализируются в выполнении запасающей, транспортной, механической и других функций. В большинстве своем паренхимные клетки имеют тонкие оболочки и разнообразную форму – изодиаметрическую или вытянутую эллипсоидальную. Протопласт клеток содержит полный набор органелл и обладает значительной пластичностью функций. Например, паренхимные клетки могут возобновлять меристематическую активность при повреждении органов, образуя каллус, который защищает места поранения. Из паренхимных клеток могут формироваться новые органы и целые растения. В определенных случаях первичная оболочка паренхимных клеток утолщается, например в запасающей ткани семян хурмы, спаржи, кофейного дерева, финиковой пальмы. В оболочках этих клеток накапливается большое количество гемицеллюлоз, играющих роль запасных веществ. В сердцевине и древесине часть паренхимных клеток одревесневает. Основные ткани развиваются из апикальных меристем и, по мере роста органов, дифференцируются и приобретают специализацию. Паренхимные клетки луба и древесины формируются вторичной латеральной меристемой камбием. Во вторичной коре – перидерме паренхимные клетки развиваются из пробкового камбия – феллогена. По выполняемой функции основная паренхима подразделяется на следующие подгруппы: ассимиляционную, запасающую, водоносную и воздухоносную. Клетки ассимиляционной паренхимы формируют хлоропласты, которые на свету осуществляют процесс фотосинтеза органических веществ. Поэтому данную ткань также называют хлоренхимой. Она развивается в органах растения, воспринимающих свет. В первую очередь к ним относятся листья или листоподобные органы, например, видоизмененные стебли. Кроме того хлоренхима развивается под прозрачной эпидермой молодых органов – стеблей, стробилов, плодов. 128 Рис. 82. Структура листа брусники на поперечном срезе: 1 – верхняя эпидерма, 2 – кутикула, 3 – палисадный мезофилл, 4 – клетки губчатого мезофилла, 5 – межклетник в губчатом мезофилле, 6 – нижняя эпидерма, 7 – устьице, 8 – склеренхима, 9 – ксилема, 10 – флоэма Хлоренхима в основном состоит из тонкостенных взаимосвязанных друг с другом клеток, между которыми имеется свободное пространство в виде межклетников (рис. 82, 83). Оно необходимо для осуществления активного газообмена в процессе фотосинтеза, включающего поглощение клетками углекислоты и выделение кислорода. Межклетники сообщаются с атмосферой через устьица в эпидерме. Клетки хлоренхимы у верхней стороны листа обычно сложены более плотно между собой и образуют столбчатую палисадную паренхиму. При ярком солнечном свете хлоропласты палисадной паренхимы располагаются у антиклинальных клеточных стенок. Слабый свет они воспринимают, перемещаясь к периклинальным стенкам. У растений умеренной зоны, осуществляющих С3-тип фотосинтеза с образованием трехуглеродных первых органических продуктов, мезофилл листа подразделяется на столбчатый и губчатый, т.е. проводящие пучки при этом окружены губчатым и столбчатым мезофиллом (рис. 82). 129 Рис. 83. Поперечный срез листа груши (Pirus): 1 – палисадная ассимиляционная паренхима, 2 – паренхимные клетки обкладки проводящих пучков, 3 – верхняя эпидерма, 4 – продолжение паренхимной обкладки проводящего пучка, 5 – волокна, 6 – ксилема, 7 – флоэма, 8 – губчатая ассимиляционная паренхима, 9 – нижняя эпидерма Растения южных широт, произрастающие в условиях жаркого и сухого климата, используют более эффективно СО2 для фотосинтеза, обладая С4-путем фотосинтеза. Эти растения, в отличие от растений умеренной зоны, открывают устьица ночью и накапливают в клетках мезофилла углекислоту в связанном виде в форме четырехуглеродной яблочной кислоты. Днем эта кислота поступает в клетки обкладки проводящих пучков, ферментативно расщепляется с выделением СО2, которая при участии света включается в обычный С3-путь фотосинтеза, характерный для растений умеренной зоны. Растения, осуществляющие С4-путь фотосинтеза, имеют следующие анатомические особенности строения. Основной объем ассимиляционной паренхимы листа у них образован губчатым и столбчатым мезофиллом, клетки которого имеют мелкие хлоропласты. В этих клетках ночью запасается углекислота. Проводящие пучки – жилки 130 Рис. 84. Электронная микрография ассимиляционной паренхимы и проводящего пучка листа злака (Сynodon dactilon), имеющего С4–путь фотосинтеза: 1 – клетка мезофилла, 2 – мелкие хлоропласты в клетках мезофилла, 3 – паренхимные клетки внутренней обкладки проводящего пучка, 4 – ксилема, 5 – паренхимная клетка наружной обкладки проводящего пучка, 6 – флоэма, 7 – межклетник, 8 – крупные хлоропласты наружной обкладки проводящего пучка листа окружены особыми клетками ассимиляционной паренхимы, называемые клетками обкладки (рис. 84). Данные клетки содержат значительно более крупные хлоропласты, осуществляющие синтез органических веществ по С3-пути фотосинтеза. Образующиеся ассимиляты поступают в проводящие ткани жилки листа. Наряду с фотосинтезом клетки хлоренхимы осуществляют запасание ассимилятов в виде первичного крахмала. В темное время суток он расщепляется ферментами на растворимые сахара, которые транспортируются из листьев к потребляющим органам. Запасающая паренхима осуществляет накопление продуктов обмена веществ – полисахаридов, белков, жиров и др. Эти ве131 щества затем расщепляются ферментами до растворимых форм и используются растением для различных видов жизнедеятельности. Эргастические вещества накапливаются в различных частях клеток запасающей паренхимы. Запасной крахмал откладывается в амилопластах, жиры – в элайопластах и цитоплазме, белки и кристаллы – в вакуолях, таннины – в цитоплазме, вакуолях или оболочках клеток. Запасающая ткань обычно сложена тонкостенными клетками, но они также могут приобретать утолщение первичных оболочек. Эта ткань развивается в значительных объемах в семенах, плодах, стеблях, луковицах, утолщенных корнях, клубнях. У многолетних растений формируется значительный объем сердцевины (рис. 85) и лучевой паренхимы – древесной и лубяной, которые служат местом резервирования (депонирования) запасных веществ. Паренхима придает древесине стойкость против инфекции и других неблагоприятных агентов. Процесс перехода светлой древесины заболони в ядровую окрашенную древесину связан с отмиранием паренхимных клеток и накоплением в них продуктов жизнедеятельности, выводимых из обмена веществ. Водоносная ткань осуществляет запасание воды. Клетки ее обычно тонкостенные, крупные, главным образом, за счет развития вакуолей. Последние содержат слизистые вещества, удерживающие воду. Такая ткань развивается в стеблях и листьях растений засушливых мест обитания – суккулентов – кактусов, агав, алоэ, молочаев и др., а также произрастающих на засоленных почвах – солерос. Воздухоносная ткань – аэренхима характеризуется развитием больших межклетников, которые разделяются слоями паренхимных клеток (см. рис. 85). Аэренхима обеспечивает газообмен тканей водных и болотных растений. Она способствует поддержанию растений во взвешенном состоянии. Такая ткань развивается в стеблях и листьях растений засушливых мест обитания – суккулентов – кактусов, агав, алоэ, молочаев и др., а также произрастающих на засоленных почвах – солерос. 132 Б А Г В Д Рис. 85. Паренхимные клетки: А, Б – паренхима сердцевины стебля березы (Betula). В молодом стебле (А) клетки имеют только первичные оболочки, а в старом (Б) также и вторичные. В, Г – паренхима аэренхимного типа (В), встречающаяся в лакунах черешков и средних жилок (Г) листьев канны (Canna). Клетки могут иметь много отростков. Д – длинная ветвистая клетка из мезофилла дисковидного цветка галлардии (Gaillardia). 1 – межклетник, 2 – межклетники ВЫДЕЛИТЕЛЬНЫЕ ТКАНИ Выделительные ткани осуществляют секрецию из растительного организма, или изолирование внутри него, продуктов обмена веществ. Эту функцию выполняют одноклеточные и многоклеточные структуры, расположенные внутри органов растения или на их поверхности. 133 Одиночные выделительные клетки, находящиеся внутри органов растения, также называются идиобластами. Чаще всего продуктами выделения служат вещества вторичного метаболизма – полифенольные соединения (дубильные вещества), терпеноиды, оксалат кальция. В зависимости от того, в каком направлении осуществляется секреция продуктов метаболизма, выделительные структуры подразделяются на внутренние и внешние выделительные структуры. Выделительные ткани встречаются во всех органах растения. Они сложены тонкостенными изодиаметрическими, либо прозенхимными клетками. Секреторная функция осуществляется за счет деятельности клеточных мембран, в особенности, комплекса Гольджи. Структуры внутренней секреции представлены отдельными клетками – идиобластами, а также вместилищами выделений: смоляными ходами, эфирно-масляными каналами, млечниками. Идиобласты накапливают таннины, кристаллы оксалата кальция, терпеноиды, слизи. В идиобластах растений из семейств лавровых, магнолиевых, перечных накапливаются терпеноиды, а в семействе мальвовых – слизи. Вместилища выделений образуются полостями, возникающими двумя основными путями – схизогенным и лизигенным. Схизогенные вместилища развиваются из межклетников, которые возникают в результате раздвигания клеток (рис. 86). Полость таких вместилищ окружается секреторным, эпителиальным слоем. Его клетки вырабатывают и выделяют во вместилище различные вещества: слизи, смолы, эфирные масла. За счет этих продуктов происходит увеличение объема вместилищ. Клетки эпителия вместилищ, как правило, окружены и метаболически взаимосвязаны с основной паренхимой органа, в котором находится вместилище. У многих хвойных растений образуются нормальные смоляные ходы в древесине. Эти растения относятся к роду сосновых: сосна, лиственница, ель, псевдотсуга, кетелеерия, катайя и дюкампопинус. Остальные сосновые – пихта, тсуга, кедр, лжелиственница и другие хвойные образуют в древесине только травматические смоляные ходы. 134 Рис. 86. Формирование смоляного вместилища в древесине сосны (Pinus) схизогенным путем. А. Дифференциация клеток эпителия. Б. Появление полости между молодыми эпителиальными клетками. В. Увеличение объема полости и числа клеток эпителия. Г. Завершение формирования смоляного вместилища. 1 – молодая клетка эпителия, 2 – клетка луча, 3 – сопровождающая паренхимная клетка, 4 – осевая трахеида ксилемы, 5 – зрелая клетка эпителия, 6 – полость вместилища Нормальный смоляной ход состоит из возникшей схизогенно полости, заполненной живицей. Эта полость выстлана клетками эпителия, которые окружены сопровождающей паренхимой. Клетки эпителия имеют более тонкую часть оболочки, обращенной к каналу, которая легко проницаема для секретируемых веществ. Обращенная наружу часть оболочки этих клеток – более толстая и практически непроницаема для смолистых веществ. Сопровождающая паренхима сложена из однородных живых клеток или состоит из мертвых клеток, расположенных несплошным слоем рядом с эпителием, и вокруг них находятся живые клетки. Сопровождающие клетки осевых смоляных ходов контактируют с лучами, получая из них продукты фотосинтеза. Эти продукты служат основой для синтеза секретируемых веществ. 135 Лизигенные вместилища формируются в результате протекания Лизигенные вместилища формипроцесса лизиса – растворения эпителиальными клетками клеток руются в результате протекания пропосле накопления ими продуктов выделения в межклетниках. цесса лизиса – растворения эпители- Рис. 87. Вместилище эфирных масел околоплодника мандарина (Citrus reticulata), полость которого возникает в результате лизиса: 1 – лизирующиеся клетки, 2 – полость Рис. 88. Членистые млечники корня одуванчика (Taraxacum officinale) в продольном разрезе: 1 – латекс, 2 – паренхима коры 136 альными клетками клеток после накопления ими продуктов выделения в межклетниках. Такого рода вместилища эфирных масел образуются, например, в околоплоднике цитрусовых (рис. 87). Млечники представляют собой вместилища, образованные клетками, накапливающими в вакуолях латекс, называемый млечным соком. Он представляет собой эмульсию белого или оранжевого цвета, содержащую таннины, терпеноиды, алкалоиды, белки, жирные масла, углеводы и т.п. Каучуконосы, например гевея бразильская из семейства молочайных, содержат каучук – производное изопрена. Существует два основных типа млечников – членистые и нечленистые. Членистые млечники образуются в результате объединения некоторого количества отдельных клеток в единую разветвленную систему (рис. 88). Они характерны для семейств маковых, сложноцветных и др. Система нечленистых млечников развивается из отдельных клеток зародыша (рис. 89). Каждая такая клетка, по мере роста растения, превращается в гигантскую клетку, которая разветвляется, проникая во все органы. Между собой такие клетки не сообщаются. Нечленистые млечники образуются у растений семейств тутовых и молочайных. Структуры внешней секреции располагаются на поверхности органов растения и связаны с покровными тканями. К ним относятся железистые Рис. 89. Нечленистые млечники олеандра (Nеrium оlеаnder). А. Формирующийся зародыш (длина 550 мкм). Молодые млечники в семядольном узле располагаются на внешней границе проводящих тканей. Начало ветвления млечника показано стрелкой (а). Б. Зрелый зародыш длиной 5 мм. Срез его шириной 75 мкм выполнен в зоне основания семядолей. Млечники распространяются от узла в семядоли и в корневую ось – гипокотиль. В. Разрастание млечника в межклетники мезофилла. 1 – семядоля, 2 – млечники трихомы и нектарники. Железистые трихомы, в свою очередь, подразделяются на головчатые волоски и железки. Железистые волоски имеют головку, в которой располагается одна или несколько секреторных клеток (рис. 90). В основании головки находится одноклеточная или многоклеточная нежелезистая ножка. Секретируемые головкой вещества, например эфирные масла у пеларгонии, накапливаются между кутикулой и оболочкой клеток. В случае разрыва кутикулы эфирное масло изливается наружу и испаряется. В последующем секреторные клетки восстанавливают слой кутикулы и накапливают под ней следующую порцию эфирного масла. У растений семейства маревых в железистых волосках накапливаются соли и вода. Железки содержат железистую многоклеточную головку, которая располагается на короткой ножке, состоящей из несекретирующих клеток. 137 Рис. 90. Трихомы секреторные. А–В – волоски листа лаванды (Lavandula vera), накапливающие эфирные масла под кутикулой секретирующих клеток (А – кутикула плотно прилегает к оболочкам к секреторным клеткам из-за отсутствия продуктов выделения, Б, В – кутикула отодвинута накапливающимся секретом от оболочек выделительных клеток). Г. Железистый волосок листа хлопчатника (Gossypium). Д. Железистый волосок с одной выделительной терминальной клеткой стебля пеларгонии (Pelargonium). Е. Клейкий волосок молодого листа груши (Pуrus). Ж. Перламутровая железка листа винограда (Vitis vinifera). 3. Жгучий волосок крапивы (Urtica urens) Железки чаще всего вырабатывают эфирные масла, что характерно для семейств губоцветных и сложноцветных. Нектарники имеют разнообразное и сложное строение. Они формируются различными частями цветка – чашелистиками, лепестками, стенкой завязи или цветоножкой. Кроме этого нектарники образуются на вегетативных органах, в этом случае они называются нецветковыми нектарниками. Нектарники имеют трубчатую, лепестковидную форму или развиваются в виде мясистых железистых разрастаний. Выделяемый нектарниками сахаристый сок привлекает насекомых и птиц, которые опыляют цветки. Этот 138 сок содержит водный раствор сахаров, белков, спиртов и ароматических веществ. Г л а в а 4. ОРГАНЫ РАСТЕНИЯ ЛИСТ ПОКРЫТОСЕМЕННОГО РАСТЕНИЯ О б ъ е к т: поперечный срез листа камелии (Сamellia japonica) Лист – орган воздушного питания растения. В нем осуществляется процесс фотосинтеза органических веществ из неорганических веществ – углекислоты воздуха и воды при участии световой энергии. Ассимиляционная паренхима или мезофилл – главная ткань листа, осуществляющая процесс фотосинтеза. Все остальные ткани листа обеспечивают необходимые условия для жизнедеятельности клеток мезофилла. Снаружи лист покрыт первичной покровной тканью – эпидермой (рис. 91). Она выполняет несколько функций. Защищает лист от механических повреждений. Предохраняет от излишнего испарения, благодаря наличию на внешней поверхности клеток эпидермы водонепроницаемого слоя кутикулы, а также воска. Эпидерма пропускает свет к мезофиллу, так как она прозрачна и не содержит пигментированных клеток, за исключением замыкающих клеток устьиц. Эпидерма обеспечивает активный газообмен между листом и атмосферой; эту функцию выполняют специализированные структуры – устьица, расположенные на нижней стороне листа. Каждое устьице имеет пору – отверстие, которое закрывается двумя замыкающими клетками. При насыщении водой клетки устьиц приобретают упругость и состояние тургора. В этом случае устьичная щель находится в открытом виде. При потере воды и увядании замыкающих клеток происходит закрывание устьичной щели. Мезофилл имеет неоднородную структуру. У верхней стороны листа он образует столбчатую паренхиму, называемую также палисадной. Здесь его клетки имеют антиклинальное удлинение (поперечное поверхности листа) и плотно прижаты друг к другу. Основная часть света поглощается палисадной паренхимой. В условиях высокого светового довольствия у листьев формируется большее число слоев клеток палисадной паренхимы, чем при низком освещении. В средней части толщи листовой пластинки и у нижней стороны расположена губчатая паренхима. Ее клетки имеют между собой 139 значительно развитую систему воздушных промежутков – межклетников. Межклетники сообщаются через устьица с атмосферой. Губ- Рис. 91. Лист камелии (Camellia japonica) на поперечном разрезе: 1 – верхняя зпидерма, 2 – столбчатая паренхима, 3 – губчатая паренхима, 4 – клетка с друзой кристаллов оксалата кальция, 5 – склереида, 6 – коллатеральный проводящий пучок, 7 – нижняя эпидерма, 8 – устьичный аппарат, 9 – ксилема, 10 – флоэма, 11 – склеренхима чатый мезофилл обеспечивает функцию газообмена между листом и атмосферой. В процессе фотосинтеза его клетки поглощают из межклетников углекислоту, поступающую через устьица, и выделяют кислород и пары воды, которые диффундируют также через устьица в атмосферу. Процесс испарения воды называется транспирацией. Водой и минеральными веществами клетки мезофилла снабжаются по ксилеме, расположенной в проводящих пучках, которые составляют густую сеть жилок листа. Ксилема содержит водопроводящие мертвые элементы – сосуды и трахеиды, а также живые паренхимные клетки, выполняющие запасающую функцию. Органические вещества – ассимиляты, образующиеся в мезофилле при фотосинтезе, транспортируются из листа по другой проводящей ткани – флоэме, также находящейся в жилках. Флоэма состоит из ситовидных трубок с клетками- спутниками, осуществляющими транспортировку веществ, а также паренхимных клеток, кото140 рые принимают участие в накоплении органических веществ и их загрузке во флоэмные окончания. Каждый проводящий пучок кроме Рис. 92. Отделительная зона листа в процессе его опадания (I) и после опадания (II): 1 – стебель побега, 2 – пазушная почка, 3 – черешок листа, 4 – эпидерма, 5 – перидерма, 6 – ксилема проводящего пучка, 7 – флоэма, 8 – отделительный слой, 9 – защитный слой, 10 – опробковевший защитный слой, 11 – закупоренное окончание проводящего пучка, 12 – проводящий пучок стебля флоэмы и ксилемы содержит на верхней и нижней стороне механическую ткань – склеренхиму, которая обеспечивает листу устойчивость в горизонтальной плоскости. Дополнительную механическую прочность листу придают отдельные клетки механической ткани – склереиды, называемые также каменистыми клетками. Эти клетки расположены в мезофилле и препятствуют его сжиманию. При старении листа начинаются процессы его отделения от стебля без повреждения живых тканей последнего, что называется опаданием. Оно сопровождается цитологическими и биохимическими изменениями клеток у основания черешка в зоне отделения (рис. 92). В этой зоне выделяются два слоя: отделительный слой, по которому происходит разлом, и защитный слой, предохраняющий 141 обнажающуюся при опадении листа поверхность от иссушения и проникновения паразитов. Отделительный слой формируется еще при образовании листа. В нем минимально развиты механические ткани, формируются укороченные проводящие элементы и преобладает паренхима. Перед опаданием листа в отделительном слое, чаще всего, начинаются клеточные деления, которые распространяются от периферии черешка к его проводящим пучкам. В проводящих элементах ксилемы начинается образование тил и отложение каллозы в ситовидных элементах и паренхимных клетках флоэмы. Эти процессы обусловливают закупоривание проводящих тканей. Главным событием при опадании листа является ферментативное разрушение клеточных оболочек. Оно приводит к утрате цементирующих свойств срединной пластинки клеточной оболочки, что связано с удалением из нее кальция. Происходит также гидролиз самих клеточных оболочек и разрыв склерифицированных проводящих элементов. Клетки отделительного слоя при этом увеличиваются в размерах, а их срединная пластинка и первичная оболочка ослизняются. Их взаимосвязь становится минимальной, и под действием собственного веса лист опадает. В обнаженных клетках защитного слоя накапливаются раневые вещества. А в межклетниках и оболочках расположенных под ними клеток откладываются защитные вещества, такие как суберин или раневая камедь. Опадание листьев задерживается фитогормоном ауксином, если его концентрация повышается до ослабляющих структурных изменений в отделительном слое. Ускорение опадания листьев происходит под действием другого фитогормона – этилена. При опадании листьев происходит синтез ферментов, разрушающих клеточную оболочку и участвующих в дыхании, в частности, пероксидазы, вызывающей усиление синтеза этилена и фосфатазы, связанной с процессами старения. ЛИСТ ГОЛОСЕМЕННОГО РАСТЕНИЯ О б ъ е к т: поперечный срез листа сосны обыкновенной (Pinus sylvesvris) Игловидный лист хвойных растений имеет складчатый мезофилл – основную ткань, осуществляющую фотосинтез органических веществ. Все остальные структуры листа обеспечивают жизнедеятельность клеток ассимиляционной паренхимы – мезофилла. 142 Снаружи лист окружен одним слоем клеток эпидермы, на поверхности которой находится кутикула (рис. 93). Эти клетки имеют утолщенные одревесневшие оболочки. Эпидерма защищает лист Рис. 93. Лист (хвоя) сосны обыкновенной (Pinus sylvestris) на поперечном разрезе: А – клеточное строение, Б – схема. 1 – эпидерма, 2 – устьичный аппарат, 3 – гиподерма, 4 – складчатая ассимиляционная паренхима, 5 – смоляной ход, 6 – эндодерма, 7 – ксилема, 8 – флоэма, 7,8 – коллатеральный проводящий пучок, 9 – склеренхима, 10 – трансфузионная ткань от механических повреждений, излишнего испарения воды, пропускает свет к мезофиллу, осуществляет активный газообмен между листом и атмосферой при помощи устьиц. Устьица состоят из околоустьичных клеток, расположенных на уровне эпидермы, и замыкающих клеток, находящихся глубже, на уровне гиподермы – второго слоя клеток листа (рис. 94). На поверхности хвои устьица ориентированы рядами, параллельными ее продольной оси. 143 Гиподерма, расположенная под эпидермой, состоит из одного, а на ребрах хвоинки – из двух-трех слоев узких длинных клеток – волокон с утолщенными одревесневшими стенками. Она придает игловидному листу механическую прочность. Рис. 94. Участок поперечного среза хвои сосны обыкновенной (Pinus sylvestris): 1 – эпидерма, 2 – гиподерма, 3 – складчатый мезофилл, 4 – ядро клетки мезофилла, 5 – хлоропласты, 6 – трансфузионная трахеида с окаймленными порами, 7 – трансфузионная паренхима с гранулами крахмала, 8 – эндодерма с гранулами крахмала, 9 – полость смоляного канала, 10 – секреторные (эпителиальные) клетки, 11 – склеренхимные клетки, 12 – подустьичная полость, 13 – околоустьичная клетка, 14 – замыкающая клетка Под гиподермой расположен складчатый мезофилл. Складки возникают вследствие углубления оболочки в полость клеток. При этом оболочка клеток приобретает увеличенную поверхность. За счет этого возрастает площадь соприкосновения протопласта клеток с апопластом – системой транспорта веществ по оболочкам кле144 ток мезофилла. Мезофилл сложен в виде плоских слоев клеток, ориентированных поперек продольной оси листа. Между ними имеются воздушные полости – межклетники, которые сообщаются через устьица с атмосферой. Во внешней части мезофилла находятся смоляные вместилища, образующиеся схизогенно, путем раздвигания межклетников. Каналы вместилищ ориентированы продольно листу. Каждый канал окружен слоем клеток эпителия с тонкими оболочками, которые вырабатывают живицу и секретируют ее в полость канала. Вокруг эпителия расположен слой клеток механической ткани – склеренхимы. Внутренняя сторона мезофилла ограничена одним слоем клеток эндодермы. Их протопласты через тангенциальные стенки осуществляют активный обмен веществ между мезофиллом и проводящим цилиндром. Благодаря избирательной проницаемости протопластов клеток эндодермы, из проводящего цилиндра в мезофилл транспортируются вода и минеральные вещества, а из мезофилла в проводящий цилиндр перемещаются ассимиляты. Радиальные стенки клеток эндодермы пропитаны суберином и лигнином и образуют пояски Каспари; они не проницаемы для воды и растворенных в ней веществ. Под эндодермой расположен проводящий цилиндр хвоинки. Его объем, в основном, заполнен трансфузионной тканью, состоящей из клеток двух типов. Около ксилемы проводящих пучков находятся пустотелые трансфузионные трахеиды с одревесневшими оболочками, имеющими окаймленные поры. Остальные клетки этой ткани – живые паренхимные тонкостенные. Трансфузионная ткань осуществляет запас веществ и их ближнюю транспортировку. В проводящем цилиндре имеются два коллатеральных проводящих пучка, содержащих ксилему и флоэму. Ксилема состоит из трахеид, которые представляют собой вытянутые мертвые клетки с окаймленными порами, осуществляющие дальнюю транспортировку воды и минеральных веществ в направлении от осевых органов к периферии. Паренхимные живые клетки ксилемы выполняют запасающую функцию. Ксилема в пучках обращена к плоской (вентральной) стороне хвоинки. Флоэма в проводящих пучках ориентирована в направлении к выпуклой (дорсальной) стороне хвоинки. Ее ситовидные живые клетки осуществляют активный перенос ассимилятов из листа в осевые органы. Паренхимные клетки флоэмы выполняют запасающую функцию. 145 Между пучками со стороны флоэмы расположена прослойка механической ткани склеренхимы. СТЕБЕЛЬ ОДНОДОЛЬНОГО РАСТЕНИЯ О б ъ е к т: поперечный срез стебля кукурузы (Zea mays) Стебель выполняет следующие функции: опорную – поддерживая в устойчивом положении надземные органы растения; транспортную – осуществляя перемещение в восходящем и нисходящем направлениях воды и растворенных в ней веществ; запасающую – накапливая воду и различные вещества. Эти функции осуществляются благодаря наличию специализированных тканей. У однодольных растений стебель сохраняет признаки первичного строения (рис. 95). Он не способен к длительному росту по диаметру после остановки продольного роста из-за отсутствия вторичных боковых образовательных тканей – камбия и феллогена. Снаружи стебель покрыт одним слоем клеток первичной покровной ткани эпидермы. Она защищает внутренние ткани от излишнего испарения воды, механических повреждений и регулирует газообмен с окружающей средой. Эпидерма формируется из наружного слоя клеток апекса побега. Под эпидермой располагается сплошной цилиндр механической ткани – склеренхимы. Клетки ее имеют толстые одревесневшие оболочки. Она придает упругость стеблю. Склеренхима и расположенные глубже ткани формируются из туники апекса побега. Внутренний объем стебля заполнен основной паренхимой. Ее крупные тонкостенные клетки выполняют запасающую функцию, а также осуществляют ближнее перемещение веществ. Основная паренхима окружает закрытые коллатеральные проводящие пучки. Они закрыты в отношении радиального роста. У однодольных растений проводящие пучки располагаются в толще стебля в несколько слоев, а у двудольных – только в один слой в виде цилиндра. В каждом проводящем пучке имеется флоэма, которая осуществляет транспортировку органических веществ в нисходящем направлении. Эти вещества образуются при фотосинтезе в надземных 146 освещенных органах. Транспортировка веществ во флоэме выполняют ситовидные трубки, деятельность которых поддерживается клетками-спутни-ками. В ситовидных трубках отсутствуют ядра клеток. Паренхимные клетки флоэмы осуществляют запасание и ближнюю транспортировку веществ. Рис. 95. Коллатеральный закрытый проводящий пучок стебля кукурузы (Zea mays): 1 – основная паренхима стебля, 2 – склеренхима, 3 – ситовидная трубка, 4 – клетка-спутник, 5 – древесинная паренхима, 6 – сетчатый сосуд метаксилемы, 7 – кольчато-спиральный сосуд протоксилемы; 8 – кольчатый сосуд протоксилемы, 9 – воздушная полость, 3, 4 – первичная флоэма, 5–9 – первичная ксилема Восходящие потоки воды и растворенных в ней минеральных веществ, поглощаемые корнем, перемещаются по сосудам и тра147 хеидам ксилемы, состоящие из отмерших пустотелых клеток. В ксилеме также имеются паренхимные клетки. Сосуды, созревающие в период раннего продольного роста стебля, называются сосудами протоксилемы, а сосуды, созревающие с период остановки продольного роста стебля – сосудами метаксилемы. Сосуды протоксилемы находятся в плоскости симметрии проводящего пучка. Рядом с ними со стороны оси стебля расположена воздушная полость. Сосуды метаксилемы – самые крупные и находятся по обе стороны от плоскости симметрии проводящего пучка. У однодольных растений в проводящих пучках имеются первичная флоэма и ксилема. Они формируются из первичной боковой меристемы прокамбия. Крупные сосуды и весь проводящий пучок окружены клетками механической ткани – склеренхимы. СТЕБЕЛЬ ТРАВЯНИСТОГО ДВУДОЛЬНОГО РАСТЕНИЯ О б ъ е к т: поперечный срез стебля древесной лианы кирказона крупнолистного (Aristolochia macrophilla) Стебель выполняет механическую функцию, проводит и запасает разнообразные вещества. У данного объекта стебель имеет переходный тип строения от пучкового к сплошному проводящему цилиндру непучкового типа. Снаружи стебель покрыт одним слоем клеток эпидермы (рис. 96). Она защищает его от механических повреждений и излишнего испарения воды. Под эпидермой находится первичная кора. Ее внешние слои образованы узкими удлиненными живыми клетками. В углах, в контакте нескольких таких клеток, расположены утолщения первичной оболочки за счет отложения целлюлозы. Эти клетки образуют механическую ткань – уголковую колленхиму. Она выполняет механическую и запасающую функции. Глубже расположена паренхима первичной коры, которая образована крупными живыми клетками с тонкими оболочками, выполняющими запасающую функцию. Самый внутренний слой клеток первичной коры, относящийся к эндодерме, образует крахмалоносные клетки. 148 Под первичной корой находится проводящий цилиндр – стела. В ее состав входит перицикл, проводящие пучки и основная ткань, расположенная в межпучковых зонах и сердцевине. В его внешней части находится механическая ткань – склеренхима перициклического происхождения (рис. 97). Она представлена сплошным цилиндром, внутренний край которого имеет ребра. Клетки склеренхимы толстостенные с одревесневшими оболочками. Рис. 96. Схема строения стебля кирказона крупнолистного (Aristolochia macrophilla): 1 – эпидерма, 2 – первичная кора, 3 – центральный цилиндр, 4 – перицикл, 5 – сердцевина, 6 – колленхима, 7 – паренхима первичной коры, 8 – эндодерма, 9 – склеренхима перицикла, 10 – паренхима перицикла, 11 – межпучковый камбий, 12 – первичная флоэма, 13 – вторичная флоэма, 14 – пучковый камбий, 15 – вторичная ксилема, 16 – первичная ксилема, 17 – сердцевинный луч 149 Рис. 97. Перицикл стебля кирказона крупнолистного (Aristolochia macrophilla): 1 – склеренхима, 2 – паренхима Под склеренхимой, между ее ребрами, в основной паренхиме расположены в один слой открытые коллатеральные проводящие пучки. Такое расположение пучков характерно для двудольных растений. Проводящие пучки открыты в отношении радиального роста, который осуществляется за счет деятельности камбия – вторичной боковой меристемы (рис. 98). Во внешнем крае пучков находятся остатки первичной флоэмы. Она формируется первичной боковой меристемой – прокамбием в продолжение продольного роста молодого стебля. Под первичной флоэмой находится вторичная флоэма, которая развивается из камбия в период после остановки продольного роста стебля. В состав флоэмы входят ситовидные трубки и клетки-спутники, осуществляющие нисходящюю транспортировку органических веществ, а также паренхимные клетки, выполняющие запасающую функцию. Под флоэмой расположен пучковый камбий. Клетки его инициального слоя остаются молодыми и способными к делению многие годы. Во внешнюю сторону инициали камбия формируют молодые клетки вторичной флоэмы, а в направлении к оси – молодые клетки вторичной ксилемы. Глубже расположена вторичная ксилема, которая формируется камбием в период после остановки продольного роста стебля. Под вторичной ксилемой на внутреннем крае проводящих пучков находится первичная ксилема. Она формируется из прокамбия во время продольного роста молодого стебля. Ксилема содержит сосуды, трахеиды и паренхимные клетки. По сосудам и трахеидам осуществляется восходящая транспортировка воды и минеральных веществ, а паренхимные клетки выполняют запасающую функцию. Между проводящими пучками в основной паренхиме находятся прослойки межпучкового камбия (см. рис. 98). В последующем они 150 формируют элементы вторичной флоэмы и вторичной ксилемы, которые заполняют пространство между пучками. В результате этого формируется сплошной проводящий цилиндр, и стебель приобретает непучковый тип строения. Внутренний объем стелы заполнен сердцевиной, сложенной паренхимными клетками, выполняющими запасающую функцию. Рис. 98. Поперечный срез центрального цилиндра стебля кирказона крупнолистного (Aristolochia macrophilla): 1 – межпучковый камбий, 2 – вторичная флоэма, 3 – пучковый камбий, 4 – вторичная ксилема, 5 – сердцевинный луч МНОГОЛЕТНИЙ СТЕБЕЛЬ ГОЛОСЕМЕННОГО РАСТЕНИЯ О б ъ е к т: поперечный срез многолетнего стебля сосны обыкновенной (Pinus sylvestris) Снаружи стебель покрыт остатками эпидермы. Под ней находится гиподерма – механическая ткань. Глубже расположена вторичная покровная ткань – перидерма (рис. 99, 100). Периферические клетки перидермы составляют проб151 ку или феллему. Феллема сложена слоями клеток, главным образом, тонкостенных, составляющих губчатую пробку. В молодых стеблях, во впадинах между поверхностными выступами, ориентированными параллельно продольной оси и видимыми на поперечных срезах (в виде лопастей), отдельными прослойками расположены каменистые клетки, образующие каменистую пробку. С возрастом в составе пробки преобладает формирование каменистых клеток. Под пробкой находится один слой клеток пробкового камбия – Рис. 99. Схема строения поперечного среза трехлетнего стебля сосны обыкновенной (Pinus sylvestris): 1 – перидерма, 2 – первичная кора, 3 – осевой смоляной канал, 4 – вторичная флоэма, 5 – камбий, 6 – вторичная ксилема, 7 – граница годичных приростов древесины, 8 – первичная ксилема, 9 – сердцевина, 10 – перимедуллярная зона, 11 – вторичный луч, 12 – первичный широкий луч с радиальным смоляным каналом, 13 – первичный узкий луч, 14 – вместилище смолы Рис. 100. Строение перидермы стебля сосны обыкновенной (Pinus sylvestris): 1 – каменистая пробка, 2 – губчатая пробка, 3 – типичная пробка, 4 – феллоген, 5 – феллодерма, 6 – лубяная паренхима, 7 – ядро клетки феллодермы, 8 – поры, 9 – клетки корки феллогена. Глубже расположена феллодерма, состоящая из живых паренхимных клеток. Перидерма окружает первичную кору, которая сложена паренхимными, тонкостенными живыми клетками, накапливающими кристаллы оксалата кальция и имеющими между собой развитую сеть межклетников. В первичной коре расположены эллипсовидные основные и округлые дополнительные смоляные вместилища. 152 Их полости выстланы эпителиальными клетками, вырабатывающими живицу. Под первичной корой находится проводящий цилиндр. Его внешняя часть образована группами отмерших облитерированных клеток первичного луба. Основной объем луба сложен вторичными элементами, сформированными камбием (рис. 101). Вторичная флоэма подразделяется на молодую проводящую, примыкающую непосредственно к камбию, и более старую, непроводящую, расположенную на удалении от камбия. Флоэма состоит: из ситовидных клеток, проводящих растворы органических веществ; белковых клеток (Страсбургера), играющих роль клеток-спутников; продольных тяжей паренхимных клеток; лучевой паренхимы; смоловыделительных эпителиальных клеток. В отличие от покрытосеменных растений, у которых ситовидные трубки и клетки-спутники имеют одну инициаль, из которой они развиваются, ситовидные клетки голосеменных имеют иные инициали, чем клетки Страсбургера. Под вторичным лубом расположен камбий, продуцирующий во внешнюю сторону молодые клетки вторичной флоэмы, а к оси стебля – молодые клетки вторичной ксилемы. Вторичная ксилема сложена в виде годичных слоев. В каждом из них на внутреннем крае имеется ранняя древесина с широкопросветными клетками, Рис. 101. Поперечный срез вторичного луба, камбиальной зоны и вторичной ксилемы стебля сосны обыкновенной (Pinus sylvestris): 1 – луч, 2 – паренхимные клетки, 3 – ситовидные клетки, 4 – ситовидные поля, 5 – крахмальные гранулы, 6 – кристалл оксалата кальция, 7 – камбий, 8 – вторичная ксилема 153 а на внешнем крае – поздняя, с толстостенными узкопросветными клетками. Древесина состоит из следующих типов клеток: 1) осевых трахеид, обеспечивающих восходящий ток воды и растворенных в ней минеральных веществ, а также выполняющих механическую функцию; 2) лучевых трахеид, осуществляющих перемещение веществ в радиальном направлении; 3) паренхимных клеток, входящих в состав лучей и сопровождающей паренхимы смоляных каналов; 4) эпителия, выстилающего полости каналов и вырабатывающего живицу. Трахеиды осевые ранние участвуют, главным образом, в транспортировке веществ, а поздние – выполняют механическую функцию. Смоляные каналы находятся, чаще всего, в поздней древесине ориентированы параллельно продольной оси стебля. Лучи, в основном, узкие однорядные. Первичные лучи пересекают весь проводящий цилиндр, а вторичные – лишь его часть. В широких многорядных лучах имеются радиальные смоляные каналы. На внутреннем крае первого годичного слоя вторичной ксилемы находятся небольшие группы клеток первичной ксилемы. С нею граничит перимедуллярная зона паренхимных клеток, примыкающих к сердцевине. МНОГОЛЕТНИЙ СТЕБЕЛЬ ДВУДОЛЬНОГО ДРЕВЕСНОГО РАСТЕНИЯ О б ъ е к т: поперечный срез многолетнего стебля липы мелколистной (Tilia cordata) Снаружи стебель покрыт остатками эпидермы. Под ней находится комплексная вторичная покровная ткань перидерма, состоящая из пробки, феллогена и феллодермы. Пробка – феллема занимает самое наружное положение в перидерме. Ее клетки – мертвые, с оболочками, пропитанными суберином. Они непроницаемы для жидких и газообразных веществ. Глубже следует феллоген – пробковый камбий. Во внешнюю сторону он вычленяет молодые клетки пробки, а к оси стебля – молодые клетки феллодермы. Феллодерма состоит из живых паренхимных клеток, выполняющих запасающую функцию. В перидерме имеются чечевички, через которые осуществляется газообмен между стеблем и атмосферой. Они представляют собой разрывы пробки, в полости 154 которых находятся тонкостенные клетки выполняющей или комплементарной ткани с межклетниками. Под перидермой находится первичная кора (рис. 102). В ее внешнем крае расположена уголковая колленхима. Клетки колленхимы живые и имеют неравномерное утолщение оболочек. Они выполняют механическую и запасающую функции и способны к фотосинтезу. Внутренняя часть первичной коры образована крупными тонкостенными паренхимными живыми клетками, выполняющими запасающую функцию. Рис. 102. Схема строения многолетнего стебля липы мелколистной (Tilia cordata): 1 – первичная кора, 2 – вторичная флоэма, 3 – вторичная ксилема, 4 – центральный цилиндр, 5 – перидерма, 6 – колленхима, 7 – паренхима первичной коры, 8 – первичные лубяные волокна, 9 – первичный луч во флоэме, 10 – вторичные лубяные волокна (твердый луб), 11 – проводящие элементы, ситовидные трубки и клетки спутники во вторичной флоэме (мягкий луб), 12 – камбиальная зона, 13 – поздняя древесина на границе годичного прироста вторичной ксилемы, 14 – первичный луч в ксилеме, 15 – вторичный луч в ксилеме, 16 – сосуд ранней древесины 155 вторичной ксилемы, 17 – первичная ксилема, 18 – перимедуллярная зона, 19 – сердцевина Первичная кора окружает проводящий цилиндр. Его наружная часть образована лубом – флоэмой. Она представлена в основном вторичными элементами. На ее внешнем крае находятся остатки первичной флоэмы. Флоэма разделена в радиальном направлении на трапецеидальные прослойки проводящего и твердого луба, которые разделены первичными лучами. Проводящий или мягкий луб сложен в виде тангенциальных слоев. Он представлен ситовидными трубками и клетками-спутниками, транспортирующими органические вещества. В мягком лубе также имеются клетки тяжевой паренхимы, образующиеся из веретеновидных клеток камбия. Между прослойками мягкого луба расположены тангенциальные слои твердого луба. Он состоит из длинных, узких, толстостенных одревесневших волокон, которые предохраняют от деформации слои проводящего луба. Лучевая паренхима сложена тонкостенными живыми клетками, которые осуществляют запасание крахмала, кристаллов оксалата кальция, а также проводят в направлениях поперек оси стебля различные вещества. Под флоэмой находится камбиальная зона. Инициальные клетки камбия в течение многих лет остаются молодыми и сохраняют способность к делению. Во внешнюю сторону они формируют молодые клетки вторичной флоэмы, а к оси стебля – вторичной ксилемы. Под камбиальной зоной расположена вторичная ксилема – древесина. Ксилема сложена в виде годичных слоев. На внутренней стороне каждого годичного слоя находится ранняя древесина, которая образуется в первой половине вегетационного периода. Она состоит из крупных клеток. Внешняя часть годичного слоя сложена поздней мелкоклеточной древесиной, формирующейся во второй половине вегетационного периода. Ксилема содержит сосуды и трахеиды, осуществляющие восходящюю транспортировку воды и минеральных веществ. Механическую роль в ней выполняют одревесневшие волокна либриформа. Паренхимные живые клетки ксилемы организованы в виде лучей, а также тяжевой древесинной паренхимы. Паренхимные клетки осу- 156 ществляют запасание различных веществ и транспортируют их на ближние расстояния в радиальном и осевом направлениях. Первичные сердцевинные лучи пересекают весь проводящий цилиндр, а вторичные, более узкие, возникают после первого года жизни стебля. На внутреннем крае самого первого годичного слоя вторичной ксилемы находятся небольшие участки первичной ксилемы, возникшие из прокамбия. В осевой части стебля расположена паренхима сердцевины, выполняющая запасающую функцию. В ее наружной стороне на границе с ксилемой, имеется перимедуллярная зона, состоящая из более мелких клеток. ПОЧКА ДРЕВЕСНОГО РАСТЕНИЯ О б ъ е к т: продольный срез вегетативной почки ели сибирской (Picea obovata) Почка представляет собой специализированную структуру побега, которая создает необходимые условия для жизнедеятельности меристематического апекса, формирующего новые почки, а также новые зачатки побегов или репродуктивных органов. Почки расположены на верхушке и на боковой части удлиненного побега ауксибласта, несущего фотосинтезирующие игловидные листья – хвою. Зрелая вегетативная почка имеет следующую структуру: 1 – меристематический апекс, 2 – зачаток побега с листовыми бугорками, 3 – зрелый укороченный расширенный стебель, несущий кроющие чешуи, которые в виде свода окружают апекс и зачаток побега (рис. 103). Ежегодно апекс осуществляет цикл органогенеза, включающий последовательное формирование фитомеров новой молодой почки, под покровами которой затем из апекса развиваются зачатки фитомеров будущего охвоенного побега с листовыми бугорками и апексами боковых осей; в репродуктивной почке апекс формирует зачаток макростробила или микростробила. (каждый фитомер состоит из листа или листоподобного образования, а также принадлежащего ему участка стебля – узла и междоузлия). В состав апекса входят следующие гистологические зоны: 1) апикальные инициальные клетки, 2) центральные материнские клетки, 3) сердцевинная меристема, 4) протодерма, 5) перифериче157 ская меристема, 6) прокамбиальные клетки (см. Гл. 3. Апексы побегов голосеменных растений). Зачаток охвоенной части побега имеет стебель, на боковой поверхности которого располагаются многочисленные листовые бугорки, представляющие собой зачатки будущих фотосинтезирующих листьев. Стебель зачатка побега имеет форму усеченного конуса. На его верхушке расположен апекс. Основание стебля располагается на расширенном укороченном стебле почки. Осевая часть зачатка стебля сложена вакуолизированными клетками сердцевинной (колончатой) меристемы, уплощенными поперек его продольной оси. Значительное их количество имеет накопление таннинов. Кроме того, в данных клетках накапливается крахмал. Рис. 103. Продольный срез почки ели сибирской (Picea obovata) в период активного заложения зачатка охвоенной части вегетативного побега новой генерации (середина августа): 1 – общий вид продольного среза верхушечной вегетативной почки, 2 – апекс на верхушке зачатка охвоенного побега, 3 – молодой зачаток 158 листового бугорка и его меристематические элементы стебля будущего охвоенного побега, недавно сформированные апексом, 4 – сердцевинная меристема зачатка охвоенной части побега, 5 – диафрагма, 6 – зачаток охвоенной части побега новой генерации, 7 – расширенный проводящий цилиндр почки, 8 – основание живой кроющей чешуи из внутреннего слоя покровов. 9 – листовые бугорки, 10 – цилиндр прокамбиальных пучков зачатка стебля, 11 – смоляное вместилище в коре почки, 12 – паренхима коры почки, 13–17 – проводящий цилиндр почки (13 – ксилема, 14 – камбий, 15 – флоэма, 16 – листовая лакуна, 17 – паренхима сердцевины), 18–23 – меристематический апекс, 18 – апикальные инициальные клетки, 19 – центральные материнские клетки, 20 – сердцевинная меристема, 21 – протодерма, 22 – периферическая меристема, 23 – прокамбиальные клетки Проводящий цилиндр зачатка побега имеет форму усеченного конуса. Он образован системой прокамбиальных пучков, которые начинаются в камбиальной зоне проводящего цилиндра стеблевой части почки и заканчиваются в листовых бугорках. Листовые бугорки расположены на боковой поверхности зачатка стебля в правильном порядке в виде нескольких листовых спиралей – парастих (см. рис. 22). Зачатки листьев плотно прижаты друг к другу, поскольку в данный период в зачатке стебля не развиваются междоузлия. Листовые бугорки, заложенные апексом самыми первыми, находятся в основании зачатка побега. Они имеют наибольший размер и степень дифференциации меристематических тканей. Это связано с тем, что после вычленения из апекса в зачатках листовых фитомеров постепенно продолжаются процессы роста, связанные с делением клеток. Наряду с этим, в них происходит формирование меристем соответствующих будущих постоянных тканей. Наиболее молодые листовые бугорки, возникшие в последнюю очередь, расположены в верхнем ярусе. Они имеют вид полусферических наименее дифференцированных бугорков. В основании зачатка побега под сердцевинной меристемой и под меристемой коры в конце вегетационного периода формируется особая отделительная ткань – диафрагма. Ее клетки образуют утолщенные оболочки. Они служат барьером между зачатком побега и зрелыми тканями почки. Как правило, ниже диафрагмы клетки сердцевины стеблевой части почки в конце вегетационного периода и зимой отмирают, образуя полость в основании зачатка побега. Диафрагма непосредственно связана с проводящими элементами ксилемы центрального цилиндра почки. Эта ткань 159 обеспечивает поддержание жизнедеятельности расположенного выше зачатка побега. Зрелый укороченный расширенный стебель почки с кроющими чешуями обеспечивает поддержание роста и жизнедеятельности меристематических структур – апекса и формируемых им зачатков, а также их защиту от неблагоприятных внешних воздействий. Стебель верхушечной почки побега чаще всего имеет больший наружный диаметр, чем стебель ниже расположенной охвоенной части побега. Это связано с тем, стебель почки имеет очень укороченные междоузлия, и его разрастание происходит в основном в радиальном направлении. Этот процесс охватывает все ткани стебля, начиная от сердцевинной паренхимы до его периферии. В результате такого роста в стебле почки формируется увеличенный диаметр сердцевины. Вокруг нее располагается система проводящих пучков центрального цилиндра, имеющая широкие листовые лакуны – промежутки, заполненные паренхимой. Из центрального цилиндра проводящие пучки следуют через паренхиму коры к основаниям кроющих чешуй почки. В проводящих пучках на внутренней, вентральной, стороне расположена флоэма, в медианной части – камбиальная зона, а на внешней, дорсальной, стороне – ксилема. Проводящий цилиндр почки непосредственно связан с проводящим цилиндром ниже расположенного стебля охвоенной части побега. Значительное развитие в почке приобретает паренхима коры, в которой развивается большое количество смоляных вместилищ. В общей сложности, стебель почки растет таким образом, что его боковая поверхность приобретает вид поверхности усеченного конуса. Причем, чем крупнее почка и чем больше она содержит слоев кроющих чешуй, тем большим становится отношение площади нижнего основание такого конуса к площади его верхнего основания. У самых крупных вегетативных почек на вершине дерева боковая поверхность их стебля даже приобретает вид воронки. Кроющие чешуи почки представляют собой видоизмененные листья – катафиллы, которые практически не способны к фотосинтезу. Их функция состоит в том, чтобы осуществлять защиту меристематических структур от неблагоприятного воздействия условий внешней среды, а также обеспечивать необходимое увеличение внутреннего свободного пространства почки для роста этих структур. Данные условия реализуются за счет того, что внешние слои кроющих чешуй почки быстро созревают и отмирают, создавая необходимый защит160 ный чехол. Это связано с тем, что в области оснований внешних слоев кроющих чешуй в стебле почки постепенно формируется перидерма, перекрывающая к ним доступ питательных веществ. Внутренние кроющие чешуи продолжительное время остаются живыми и способными к интеркалярному, вставочному, росту. Эти слои кроющих чешуй сохраняют взаимосвязь с живыми тканями стебля. КОРЕНЬ Корень представляет собой орган почвенного питания растения. Он выполняет следующие основные функции: поглощение веществ из почвы, их трансформацию, накопление и транспорт, а также механическое закрепление растения в почве. По продольной оси молодой корень имеет следующие морфологические и функциональные элементы (рис. 104). На кончике корня расположен корневой чехлик, который покрывает апикальную меристему корня. Он предохраняет ее от повреждения и способствует продвижению растущего корня между частицами почвы. На верхушке апикальной меристемы находятся инициальные клетки. Они продуцируют меристематические клетки, из которых развиваются корневой чехлик, ризодерма, кора и центральный цилиндр. Выше инициальных клеток апекса следует зона деления, обозначаемая также как меристема, в которой клетки интенсивно делятся. Клетки меристемы сложены продольными рядами. Каждый ряд берет свое начало от инициальной клетки ряда. В пределах зоны деления клетки в ряду сохраняют небольшие размеры (например, у проростков пшеницы – 11–22 мкм). Над меристемой расположена зона растяжения, клетки которой прекращают деление и активно растут. По мере прекращения клеточных делений, в зоне растяжения клетки в продольных рядах увеличиваются в длину в несколько раз (например, у проростков пшеницы – более чем в 10 раз, достигая длины 280 мкм). Далее по оси следует зона корневых волосков, которые обеспечивают активное поглощение корнем веществ из почвы. В этой зоне корня часть клеток ризодермы, называемых трихобластами, образует цилиндрические выросты в виде корневых волосков. Они много- 161 кратно увеличивают всасывающую поверхность между корнем и почвенным раствором. Корень однодольного растения О б ъ е к т: поперечный срез корня ириса (Iris germanica) Корни большинства однодольных растений сохраняют признаки первичного строения и не обладают вторичным радиальном ростом из-за отсутствия способности формировать вторичные боковые меристемы – камбий и феллоген. На поперечном срезе корня видны следующие ткани (рис. 105). Поверхность корня покрыта одним слоем клеток ризодермы – первичной покровной ткани. Часть ее клеток, называемых трихобластами, в зоне всасывания образуют выросты в виде тонкостенных корневых волосков. Корневые волоски – трихомы осуществляют активное избирательное поглощение из почвы разнообразных веществ и воды. Этот процесс связан с затратой энергии в виде АТФ, которая синтезируется в результате дыхания клеток. Под ризодермой находится широкая первичная кора. В корне кора занимает значительно больший относительный объем, чем в стебле. Кора корня подразделяется на три коцентрически расположенные зоны, включающие внешнюю – экзодерму, среднюю, самую широкую – мезодерму, и внутреннюю – эндодерму. 162 Рис. 104. Корень проростка пшеницы (Triticum aestivum): А – общая схема строения, Б – наружные слои клеток различных зон. 1 – корневой чехлик; 2 – инициальные клетки корневого чехлика – калиптроген; 3 – меристема; 4 – зона растяжения клеток; 5 – зона корневых волосков; 6 – окончание зоны живых корневых волосков; 7 – корневые волоски; 8 – начало роста корневого волоска из клетки ризодермы – трихобласта Рис. 105. Поперечный срез корня ириса касатика (Iris germanica) в зоне проведения веществ: 1 – ризодерма, 2 – трехслойная экзодерма, 3 – мезодерма, 4 – эндодерма (2-4 – первичная кора), 5 – пропускная клетка эндодермы, 6 – перицикл, 7 – первичная ксилема, 8 – первичная флоэма, 9 – механическая ткань (6–9 – центральный цилиндр) Экзодерма сложена двумя-тремя слоями паренхимных клеток. В зоне, расположенной выше живых корневых волосков, ее клетки пробковеют и переходят к выполнению защитной функции. Основной объем коры составляет паренхима мезодермы. Экзодерма и мезодерма осуществляют накопление веществ и транспортируют их в радиальном направлении к внутреннему слою коры – эндодерме и к внешнему краю коры – ризодерме. Большинство клеток эндодермы имеет утолщенные оболочки. Их радиальные стенки не проницаемы для растворов, поскольку они пропитаны суберином. На тангенциальном срезе радиальные стенки клеток эндодермы имеют вид опробковевших многогранников, называемых также поясками Каспари. Часть клеток эндодермы имеет тонкие оболочки. Эти клетки называются пропускными. Через свои протопласты они осуществляют активный избирательный перенос веществ между корой и проводящим цилиндром. Из коры в проводящий цилиндр пропускные клетки проводят воду и минеральные вещества, а из проводящего цилиндра в кору они транспортируют органические вещества, которые обеспечивают поддержание процессов поглощения веществ корнем. Наружный слой проводящего цилиндра состоит из клеток перицикла, часть которых способна возобновлять пролиферацию. Его клетки совместно с клетками прилегающих тканей осуществляют заложение боковых корней. 163 Под перициклом расположена проводящая система корня, имеющая полиархный тип организации (ксилема сложена в виде нескольких лучей – более четырех-пяти, видимых на поперечном срезе). Проводящая система корня представлена единственным проводящим пучком радиального типа. В этом пучке по оси расположена первичная ксилема. Она выглядит в виде ребристого цилиндра. На периферии, на границе с перициклом, расположены узкопросветные сосуды – протоксилемы, возникающие первыми в корне. Они имеют спиральные и кольчатые утолщения вторичной оболочки. Внутрь от них располагаются более крупные сосуды – метаксилемы, которые образуются позже, причем каждый последующий сосуд метаксилемы формируется ближе к оси. Ближе всего к оси располагаются самые широкопросветные, обычно пористые, сосуды метаксилемы, развивающиеся наиболее поздно. В самой осевой части корня находится механическая ткань с равномерно утолщенными одревесневшими оболочками. Присутствие сердцевины в корне – явление нетипичное, а если она и развивается, то значительно уступает по размерам сердцевине стебля. В углублениях между ребрами первичной ксилемы находятся продольные тяжи первичной флоэмы. Она состоит из ситовидных трубок, клеток-спутников и паренхимных клеток. Самые ранние элементы протофлоэмы находятся около перицикла, а более крупные клетки метафлоэмы – ближе к оси корня. Тяжи флоэмы отделены от ксилемы прослойкой живых тонкостенных клеток осевой паренхимы. Корень двудольного травянистого растения О б ъ е к т: корень тыквы (Cucurbita pepo) У двудольных растений корень сохраняет первичное строение в зоне всасывания веществ, где он имеет сходные черты с корнем однодольных растений. Однако выше по оси, в зоне проведения веществ, у корня двудольного растения возникают вторичные изменения (рис. 106). Они состоят в том, что между первичной ксилемой и первичной флоэмой в радиальном проводящем пучке закладывается вторичная латеральная меристема – камбий (рис. 164 107). Его инициальные клетки начинают продуцировать в направлении к оси корня новые клетки вторичной ксилемы, а в направлении к периферии корня – новые клетки вторичной флоэмы. В результате активного нарастания вторичных проводящих тканей ксилемы и флоэмы корень приобретает значительную толщину. Очень часто в корнеплодах эти ткани служат местом отложения запасных питательных веществ. Наряду с деятельностью камбия активизируется пролиферация (деление клеток) перицикла, который начинает формировать элементы перидермы: во внешнюю сторону – молодые клетки пробки, а к оси корня – единичные слои клеток феллодермы. Созревающие клетки пробки изолируют расположенные снаружи от нее ризодерму и первичную кору, что приводит к отмиранию последних. Вследствие этого в корнях вторичного строения между перидермой и проводящим цилиндром отсутствует первичная кора, которая характерна для побегов двудольных растений. Корень вторичного строения двудольного растения тыквы имеет следующие ткани (см. рис. 107). Его поверхность покрывает пробка, состоящая из нескольких слоев омертвевших клеток, оболочки которых пропитаны суберином. Под нею находится феллоген – вторичная латеральная меристема, продуцирующая перидерму. Феллоген расположен над феллодермой, состоящей из нескольких слоев живых паренхимных клеток, формируемых феллогеном. Пробка, феллоген и феллодерма образуют комплекс вторичной покровной ткани – перидерму. Перидерма окружает проводящий цилиндр корня. Он состоит из нескольких проводящих пучков, разделенных между собой широкими лучами. Во внешней стороне пучков расположен луб. Его основной объем составляет вторичная флоэма. В ее наружном крае находятся узкие тангенциальные прослойки первичной флоэмы, сформированные прокамбием. За вторичной флоэмой к оси корня следует пучковый камбий. Из проводящих пучков камбий продолжается в тангенциальном направлении в лучах, образуя сплошную цилиндрическую прослойку. Во внешнюю сторону камбий формирует новые 165 слои клеток вторичной флоэмы и радиальных лучей. В сторону оси он продуцирует новые элементы вторичной Рис. 106. Схема изменений, происходящих в корне двудольных растений при переходе от первичного строения к вторичному: 1 – эпиблема, 2 – первичная кора, 3 – первичная флоэма, 4 – камбий, 5 – первичная ксилема, 6 – эндодерма, 7 – перицикл, 8 – вторичная ксилема, 9 – вторичная флоэма, 10 – радиальный луч, 11 – паренхима вторичной коры, 12 – перидерма Рис. 107. Вторичное строение корня тыквы (Сucurbita реро): 1 – пробка, 2 – основная паренхима, 3 – радиальный луч, 4 – первичная флоэма, 5 – вторичная флоэма, 6 – пучковый камбий, 7 – вторичная ксилема, 8 – межпучковый камбий, 9 – первичная ксилема Ряд растений развивает большой объем главного корня, образуя корнеплоды. Эти органы, помимо выполнения обычных функций 166 корня, осуществляют также отложение значительных количеств запасных веществ. Дополнительный объем корнеплодов формируется за счет значительного развития флоэмы, например, у моркови и петрушки, ксилемы – у редьки, турнепса. Корнеплод свеклы формирует увеличенный объем за счет развития добавочных концентрических слоев камбия, который, в свою очередь, дает начало множеству дополнительных коллатеральных проводящих пучков, окруженных паренхимой. В конечном итоге, большой объем корнеплодов, главным образом, формируется за счет развития в них паренхимной ткани. Примером растения, осуществляющего накопление запасных питательных веществ в корне, может служить морковь – представитель семейства зонтичных (рис. 108). В корне моркови вторичного строения имеются следующие ткани. На периферии расположены пробка, феллоген и феллодерма – элементы перидермы. Под нею Рис. 108. Корень вторичного строения моркови (Daucus carota var. Sativa): 1 – пробка; 2 – паренхима вторичной коры, 3 – камбий, 4 – вторичная ксилема, 5 – первичная ксилема, 6 – радиальный луч, 7 – первичная и вторичная флоэма находится паренхима вторичной коры. Далее к оси следует проводящий цилиндр. На его внешнем крае располагаются остатки пер167 вичной флоэмы, подстилаемые достаточно широкой вторичной флоэмой. Она содержит ситовидные трубки с клетками-спутниками и сильно развитые радиальную и осевую системы паренхимных клеток, которые выполняют запасающую функцию. Под вторичной флоэмой расположена камбиальная зона, которая находится на внешней границе двух секторов вторичной ксилемы. Ксилема содержит сосуды, трахеиды, клетки осевой и радиальной паренхим. Между секторами проводящих элементов ксилемы расположены два сектора лучей паренхимы, проходящих через камбий во вторичную флоэму. В осевой части корня находится диархная первичная ксилема, которая имеет две группы сосудов в виде лучей, расположенных под углом 180 о . Корни травянистых двудольных растений, также как и многолетние корни хвойных растений, имеют небольшое количество лучей первичной ксилемы – от двух до четырех. Корень многолетнего двудольного древесного растения О б ъ е к т: поперечный срез корня липы мелколистной (Tilia cordata) У многолетних двудольных древесных растений, в отличие от травянистых однолетних растений, в корне формируются годичные слои вторичной ксилемы и вторичной флоэмы. Кроме того, большое развитие получает перидерма, годичные слои которой в старых корнях преобразуются в третичную покровную ткань – корку. По этим признакам многолетний корень сходен со структурой многолетнего стебля. Однако корень имеет целый ряд особенностей строения, отличающих его от стебля. К ним относятся наличие по оси корня на поперечном срезе многолучевой фигуры первичной ксилемы – 5–7 лучей (полиархный тип) и отсутствие сердцевины. Для корня характерны также менее выраженная годичная слоистость ксилемы и большее развитие в ней паренхимных клеток, выполняющих запасающую функцию. Во вторичной флоэме и ксилеме корня менее развита склеренхима в виде лубяных волокон и либриформа. В многолетнем корне отсутствует паренхима первичной коры. 168 Многолетний корень липы имеет следующее гистологическое строение (рис. 109). Снаружи он покрыт пробкой, за которой глубже следуют феллоген и тонкий слой феллодермы, составляющие элементы перидермы. Под перидермой находится проводящий цилиндр корня. На его внешней стороне расположена флоэма. Она включает следующие типы клеток: ситовидные трубки, клетки-спутники, осевую паренхиму, паренхиму лучей, клетки механической ткани – лубяные волокна. Основной объем луба занимает вторичная флоэма. Небольшие остатки первичной флоэмы находятся на верхушках трапециевидных прослоек вторичного проводящего и твердого луба. Проводящий луб включает ситовидные трубки и клетки-спутники, твердый луб представлен лубяными волокнами. Эти трапециевидные прослойки разделены широкими первичными лучами. Глубже расположена камбиальная зона. Под ней находится широкая вторичная ксилема, сложенная концентрическими слоями годичных приростов. 169 Рис. 109. Схема строения многолетнего корня липы (Tilia cordata) на поперечном срезе: 1 – первичная ксилема, 2 – вторичная ксилема, 3 – камбий, 4 – вторичная флоэма, 5 – первичная флоэма, 6 – перидерма, 7 – пробка, 8 – феллоген, 9 – феллодерма, 10 – остатки проводящих элементов первичной флоэмы, 11 – паренхима луча во вторичной флоэме, 12 – друза кристаллов оксалата кальция в клетке луча, 13 – лубяные волокна (твердый луб) во вторичной флоэме, 14 – проводящие элементы вторичной флоэмы (ситовидные трубки и клетки-спутники), 15 – вторичный луч во вторичной флоэме, 16 – граница годичного прироста вторичной ксилемы, 17 – сосуды поздней древесины, 18 – сосуды ранней древесины, 19 – первичный луч во вторичной ксилеме, 20 – сосуды первичной ксилемы, 21 – паренхима первичной ксилемы В состав ксилемы входят сосуды, трахеиды, волокна либриформа, осевая и радиальная паренхимы. Осевую часть корня занимает многолучевая фигура первичной ксилемы, состоящая из сосудов протоксилемы и метаксилемы. В осевой части первичной ксилемы находится паренхима ксилемы. Корень многолетнего хвойного древесного растения О б ъ е к т: поперечный срез корня сосны обыкновенной (Pinus sylvestris) Многолетний корень хвойного растения имеет ряд общих признаков строения, характерных для корней древесных растений (рис. 110). К ним относятся более значительное, по сравнению со стеблем, развитие луба, лубо-древесинные лучи шире, менее четко выражены границы годичных приростов древесины. Трахеиды на поперечном сечении крупнее и имеют более тонкие оболочки, поэтому древесина корней механически менее прочная, чем у стеблей. Во флоэме и ксилеме корня более развита паренхима, выполняющая запасающую функцию. 170 Рис. 110. Схема строения корня сосны обыкновенной (Pinus sylvestris) на поперечном срезе: 1 – перидерма, 2 – феллодерма, 3 – луч, 4 – вторичная флоэма (вторичный луб), 5 – осевой смоляной канал, 6 – воздухоносная полость, 7 – первичная ксилема, 8 – камбий, 9 – вторичная ксилема, 10 – первичный смоляной канал Снаружи многолетний корень сосны покрыт вторичной покровной тканью – перидермой. Основную покровную функцию в ней выполняют слои мертвых клеток пробки, занимающие периферическое положение. В ее состав входят клетки обычной пробки, губчатой пробки с тонкими оболочками и каменистой пробки, представленной в виде отдельных клеток-филлоидов с сильно утолщенными одревесневшими слоистыми и пористыми оболочками. Новые клетки пробки формируются феллогеном – пробковым камбием, подстилающим пробку. Из феллогена также развиваются в направлении к оси корня крупные крахмалоносные паренхимные клетки феллодермы. Она сложена в три-четыре слоя клеток. Феллодерма окружает проводящий цилиндр корня. Значительный объем в нем занимает расположенный периферически луб. Его внешняя часть относится к непроводящему лубу, который сложен старыми слоями клеток. Многие из них сдавлены, а ситовидные клетки облитерированы. Здесь также имеются воздухоносные полости, возникшие рексигенно в результате разрыва клеток. Ближе к камбию находится проводящая часть луба. Его ситовидные клетки ориентированы правильными радиальными рядами. Между ними находятся клетки лучевой и осевой – тяжевой паренхимы. Последняя образует однорядные тангенциальные полоски. Клетки лубяной паренхимы заполнены крахмалом, каплями жира, смолы и одиноч171 ными кристаллами оксалата кальция. В этих клетках много дубильных веществ. На внутренней границе луба находится камбиальная зона. В ней снаружи от инициального слоя камбия расположены новые клетки вторичной флоэмы, а внутрь от него – новые клетки вторичной ксилемы. Вторичная ксилема сложена в виде концентрических слоев годичных приростов древесины, неравномерно утолщенных по периметру. По сравнению с ксилемой стебля, трахеиды в корне имеют большую площадь поперечного сечения и сравнительно тонкие оболочки. Они расположены правильными радиальными рядами. В трахеидах содержатся крупные окаймленные поры, которые, в основном, размещены на радиальных стенках в два и даже в три ряда, что характерно только для корней. У стеблевой древесины трахеиды образуют, в основном, одиночные ряды крупных окаймленных пор. Каждый слой годичного прироста древесины корня снаружи ограничен одним-двумя слоями более толстостенных поздних трахеид, которые сжаты в радиальном направлении. Они в большей мере выполняют механическую функцию и имеют немного пор. В поздней части годичных слоев древесины расположены крупные осевые смоляные каналы, одиночные или сгруппированные в тангенциальном направлении по два и более. Полость смоляных каналов окружена тонкостенными эпителиальными клетками, которые окружены массивными обкладками сопровождающей паренхимы. Вторичная ксилема пересечена в радиальном направлении многочисленными одно-двухрядными лучами, расходящимися в стороны от первичной ксилемы. Клетки лучевой паренхимы запасают крахмал. В широких лучах содержатся радиальные смоляные каналы. В осевой части корня находится цепочка трахеид диархной (имеющей на поперечном срезе два луча) первичной ксилемы. На ее концах расположены трахеиды протоксилемы, за которыми следуют два первичных осевых смоляных канала перициклического происхождения. Обкладка сопровождающей паренхимы этих каналов объединена с паренхимой первичной ксилемы. Микориза, корневые клубеньки Снабжение растения водой и питательными веществами корнем усиливается, благодаря его симбиотической ассоциации со специфическими грибами, называемой микоризой. Данный вид 172 симбиоза широко распространен в природе. С анатомической точки зрения, различают эктотрофные и эндотрофные микоризы. Эктотрофные микоризы почти исключительно образуются грибами базидиомицетами и распространены, преимущественно, у лесных деревьев среди видов семейств сосновых, ивовых, березовых, буковых, ореховых, цезальпиниевых, липовых. Эндотрофные микоризы образуются грибными симбионтами – фикомицетами и распространены, больше всего, у травянистых растений, хотя и встречаются у древесных (плодовых деревьев, ясеня, тюльпанного дерева). Существуют также эктоэндотрофные микоризы, для которых свойственно проникновение отдельных гиф в клетки корня. Эктотрофная микориза характеризуется образованием плотного чехла из плектенхимы (грибной ткани) или гифальной мантии на молодых неопробковевших окончаниях корней и внедрением гиф между клетками коры корня (рис. 111). Клетки корня при этом не обнаруживают заметных патологических симптомов. Внедряясь между клетками первичной коры, гриб разделяет их между собой, образуя 173 Рис. 111. Строение корней эвкалипта (Eucalyptus) неинфицированного (III, I) и инфицированного (IV, II) микоризным грибом. Продольные осевые срезы корней: I – неинфицированного, II – микоризного. Поперечные срезы полностью дифференцированной зоны корня: III – неинфицированного, IV – микоризного. 1 – корневой чехлик, 2 – меристема, 3 – гифальный чехол или мантия, 4 – сеть Гартига, 5 – утолщенные оболочки внутренней первичной коры, 6 – ризодерма, 7 – наружная первичная кора, 8 – внутренняя первичная кора, 9 – эндодерма (заштрихованные участки пропитаны таннином), 10 – корневой волосок, 11 – одревесневшая протоксилема, 12 – сплющенные остатки клеток чехлика сеть гифов, называемую сетью Гартига. Образование микоризы приводит к подавлению развития корневых волосков и уменьшению объема апикальной меристемы и корневого чехлика. Зона дифференциации у микоризных корней расположена значительно ближе к апексу, по сравнению с неинфицированными корнями. Для микоризных корней характерны утолщение внутренней первичной коры и радиальное удлинение клеток ризодермы. Эктомикоризные корни контактируют с почвой через грибной мицелий. Большая часть питательных веществ, в том числе азота и фосфора, поступает в дерево через микоризы. Считается, что растение-хозяин секретирует сахара, аминокислоты и другие органические вещества, делая их доступными для гриба. Положительное влияние микоризы на рост деревьев, особенно на почвах, бедных питательными веществами, объясняется улучшением питания при помощи гриба, улучшением азотного питания в результате расщепления гумуса грибом, активирования обмена веществ в дереве, благодаря снабжению его ростовыми веществами, выделяемыми микоризными грибами. К растениям, испытывающим большую потребность в микоризе, относятся большинство хвойных, дуб, бук, граб. Эндотрофная микориза широко распространена в природе, но исследована она гораздо меньше, чем эктотрофная. При этом симбиозе гриб не образует заметной мантии, а внедряется внутрь кле174 ток корня. Эндомикоризные корни отличаются более темной окраской по сравнению с неинфицированными корнями. Ассоциация корней с азотфиксирующими бактериями видов Rhizobium приводит к развитию корневых клубеньков, благодаря которым обеспечивается снабжения растения-хозяина азотом. Клубеньки образуются на корнях многих бобовых растений. Ряд древесных растений, например ольха, облепиха, лох узколистный и др., Вступают в симбиотическое сожительство с актиномицетами (лучистыми грибами), в результате чего эти растения становятся способными усваивать газообразный азот. У них из первоначально маленьких клубеньков в последующем образуются одревесневшие долголетние азотфиксирующие ризотамнии, которые, в свою очередь, образуют плотные скопления на корнях. В настоящей главе рассмотрена структура вегетативных органов растений с наиболее типичным расположением тканей, как правило, на поперечных сечениях, проходящих по середине органов – стебля или корня. Однако, например, в переходной зоне контакта стебля с корнем, особенно на ранних этапах развития проростка, обнаруживается специфическая структура проводящей системы центрального цилиндра. Она заключается в том, что цилиндрическая система проводящих тканей корня, не имеющего сердцевины, и система проводящих пучков с сердцевиной верхнего гипокотиля сложным образом соединяются между собой в переходной зоне. Таким образом, характеризуя гистологическую структуру органов растения, следует учитывать ее видоизменение во взаимных контактах органов, на краях органов и т.п. Особый интерес представляет вопрос о трансформации гистологической структуры органов под действием факторов окружающей среды – биотических и абиотических, что служит предметом исследования специальных разделов анатомии растений – экологической анатомии, патологической анатомии и т.п. 175 Библиографический список Основная литература 1. Эсау К. Анатомия семенных растений. Кн. 1 и 2. М.: Мир, 1980. 2. Эсау К. Анатомия растений. М.: Мир, 1969. 564 с. 3. Родионова А.С., Барчукова М.В. Ботаника. Л.: Агропромиздат, 1990. 303 с. 4. Васильев А.Е., Воронин Н.С., Еленевский А.Г., Серебрякова Т.И. Ботаника. Морфология и анатомия растений. М.: Просвещение, 1988. 478 с. 5. Яценко-Хмелевский А.А. Краткий курс анатомии растений. М.: Просвещение, 1961. 282 с. Дополнительная литература 1. Рейнвн П., Эверт Р., Айкхорн С. Современная ботаника. Кн. 1 и 2. М.: Мир, 1990. 344 с. 2. Лотова Л.И. Анатомия коры хвойных. М.: Наука, 1987. 152 с. 3. Лотова Л.И. Микроструктура коры основных лесообразующих лиственных деревьев и кустарников восточной Европы. М.: КМК Scientific Press, 1988. 113 c. 4. Лотова Л.И. Морфология и анатомия высших растений. М.: Эдиториал УРСС, 2000. 528 с. 5. Яковлев Г.П., Аверьянов Л.В. Ботаника для учителя. Ч. 1. М. Просвещение, 1996. 220 с. 176 6. Яковлев Г.П., Аверьянов Л.В. Ботаника для учителя. Ч. 2. М. Просвещение. 1997. 7. Яценко-Хмелевский А.А., Барчукова М.М., Кобак К.И. Анатомия растений. Побег и корень: Лабораторный практикум. Л.: ЛТА, 1982. 63 с. 8. Яценко-Хмелевский А.А., Барчукова М.М., Кобак К.И. Анатомия растений. Строение древесины: Лабораторный практикум. Л.: Изд. ЛТА. 1982. 9. Хржановский В.Г., Пономаренко С.Ф. Практикум по курсу общей ботаники. М.: Высшая школа, 1979. 422 с. 10. Практикум по анатомии растений: учебное пособие для студентов биологических спец. вузов / Барыкина Р.П., Кострикова Л.Н., Кочемарова И.П. и др.; Под ред. Транковского Д.А. 3-е изд., перераб. и доп. М.: Высш. шк., 1979. 224 с. 11. Чавчавадзе Е.С. Древесина хвойных. Морфологические особенности, диагностическое значение. Л., 1979. 192 с. 12. Еремин В.М. Атлас анатомического строения коры сосновых СССР. М., 1978. 203 с. 13. Атлас ультраструктуры растительных тканей. Петрозаводск: Карелия, 1980. 456 с. 14. Киселева Н.С., Шелухин Н.В. Атлас по анатомии растений. Минск: Вышэйш. шк., 1969. 288 с. Оглавление П р е д и с л о в и е............................................................................................ 3 Глава 1. Введение .................................................................................. 4 Краткий обзор анатомической техники и методов подготовки объектов к исследованию .................................................................... 6 Настройка просвечивающего светового микроскопа ........................ 9 Классификация тканей растений ...................................................... 11 Развитие и общая структура растительного организма .................. 11 Глава 2. Клетка...................................................................................... 16 Мембраны клетки ................................................................................... 18 Цитоплазма ............................................................................................. 20 Ядро ........................................................................................................ 20 Пластиды ................................................................................................ 22 Митохондрии........................................................................................... 23 Микротельца – пероксисомы, цитосомы .............................................. 23 Вакуоли ................................................................................................... 24 Структуры, связанные с плазмалеммой. .............................................. 24 Рибосомы. ............................................................................................... 25 Аппарат Гольджи .................................................................................... 25 Эндоплазматический ретикулум ........................................................... 25 Глобулы липидов.................................................................................... 26 Микротрубочки........................................................................................ 26 177 Микрофиламенты ................................................................................... 27 Эргастические вещества. ...................................................................... 27 Оболочка клетки ................................................................................. 30 Структура и состав клеточной оболочки .......................................... 31 Формирование клеточной оболочки ................................................. 36 Поры клеточной оболочки ................................................................. 39 Межклетники....................................................................................... 42 Глава 3. Ткани ....................................................................................... 43 Образовательные ткани ........................................................................ 43 Апикальная меристема побега ........................................................... 44 Апексы побегов голосеменных растений ....................................... 45 Апексы побегов покрытосеменных растений ................................. 52 Маргинальная меристема ............................................................... 53 Апикальная меристема корня ............................................................ 54 Заложение бокового корня .............................................................. 59 Латеральные меристемы.................................................................... 64 Первичные латеральные меристемы ............................................ 64 Прокамбий .................................................................................... 64 Перицикл ...................................................................................... 65 Вторичные латеральные меристемы ............................................... 65 Камбий .......................................................................................... 65 Феллоген ...................................................................................... 70 Интеркалярные меристемы ............................................................... 71 Раневые меристемы .......................................................................... 72 Покровные ткани .................................................................................... 73 Эпидерма ....................................................................................... 73 Трихомы.......................................................................................... 77 Корневые волоски ......................................................................... 78 Перидерма .......................................................................................... 80 Корка ................................................................................................... 81 Проводящие ткани ................................................................................. 82 Флоэма .................................................................................................... 82 Первичная флоэма ........................................................................ 89 Вторичная флоэма......................................................................... 90 Ксилема .............................................................................................. 92 Трахеальные элементы ................................................................. 94 Волокна........................................................................................... 98 Паренхимные элементы ................................................................ 99 Проводящие пучки ....................................................................... 104 Строение древесины ................................................................... 105 Древесина хвойных растений ......................................................... 106 Древесина сосны обыкновенной................................................. 107 Древесина лиственницы.............................................................. 111 Древесина покрытосеменных растений ......................................... 115 Древесина липы мелколистной .................................................. 115 Древесина дуба черешчатого ..................................................... 118 Механические ткани ............................................................................. 121 178 Колленхима ...................................................................................... 122 Склеренхима ................................................................................... 123 Волокна......................................................................................... 124 Склереиды .................................................................................... 126 Основные ткани .................................................................................... 128 Выделительные ткани .......................................................................... 133 Глава 4. Органы растения ................................................................ 139 Лист покрытосеменного растения. .................................................. 139 Лист голосеменного растения ......................................................... 142 Стебель однодольного растения .................................................... 146 Стебель травянистого двудольного растения ............................... 148 Многолетний стебель голосеменного растения ............................ 151 Многолетний стебель двудольного древесного растения ........... 154 Почка древесного растения ............................................................ 157 Корень .......................................................................................... 160 Корень однодольного растения ................................................. 161 Корень двудольного травянистого растения ............................. 164 Корень многолетнего двудольного древесного растения ........ 168 Корень многолетнего хвойного древесного растения .............. 170 Микориза, корневые клубеньки ................................................... 172 Б и б л и о г р а ф и ч е с к и й с п и с о к ..................................................... 176 ДЛЯ ЗАМЕТОК 179 Владимир Борисович Скупченко АНАТОМИЯ РАСТЕНИЙ Учебное пособие для студентов специальностей «Лесное хозяйство» и «Садово-парковое и ландшафтное строительство» направления «Лесное хозяйство и ландшафтное строительство» Редактор и техн. редактор – Н.А. Теплякова Компьютерная верстка – В.Ф. Сергеев Подписано в печать с оригинал-макета 11.05.04. Формат 60х84/16. Бумага офсетная. Печать трафаретная. Уч.-изд. л. 11,25. Печ. л. 11,25. Тираж 300 экз. Заказ 140. С 20. Санкт-Петербургская государственная лесотехническая академия Издательско-полиграфический отдел СПбГЛТА 194021, Санкт-Петербург, Институтский пер., 3 180